
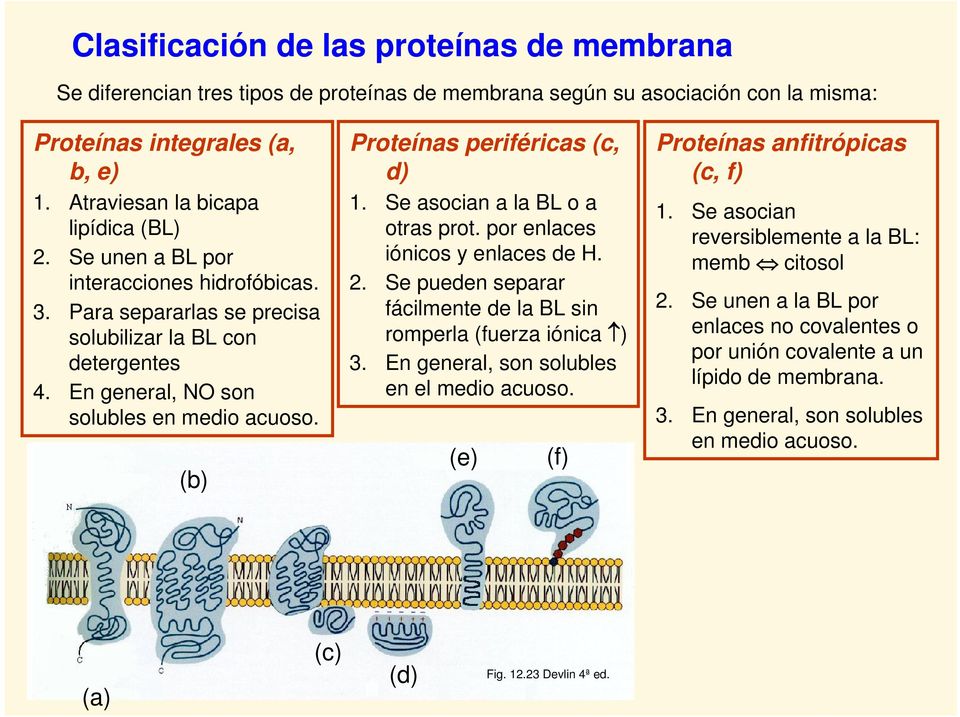
Formas de vida. Un ser vivo u organismo es un conjunto material de organización compleja, en la que intervienen sistemas de comunicación molecular que lo relacionan internamente y con el medio ambiente en un intercambio de materia y energía de una forma ordenada, teniendo la capacidad de desempeñar las funciones básicas de la vida que son la nutrición, la relación y la reproducción, de tal manera que los seres vivos funcionan por sí mismos sin perder su nivel estructural hasta su muerte.

La reproducción es una característica básica de los seres vivos. En la parte superior de la figura se aprecia una bacteria reproduciéndose por fisión binaria.
Resulta fácil, habitualmente, decidir si algo está vivo o no. Ello es debido a que los seres vivos comparten muchos atributos. Asimismo, la vida puede definirse según estas propiedades básicas de los seres vivos, que nos permiten diferenciarlos de la materia inerte:
- Organización. Las unidades básicas de un organismo son las células. Un organismo puede estar compuesto de una sola célula (unicelular) o por muchas (pluricelular).
- Homeostasis. Los organismos mantienen un equilibrio interno, por ejemplo, controlan activamente su presión osmótica y la concentración de electrolitos.
- Relación o Irritabilidad. Es una reacción ante estímulos externos y permite a los seres vivos detectar u obtener información del medio en el que viven, tomar las decisiones acertadas y elaborar una respuesta adecuada para su supervivencia. Una respuesta puede ser de muchas formas, por ejemplo, la contracción de un organismo unicelular cuando es tocado o las reacciones complejas que implican los sentidos en los animales superiores.
- Metabolismo. Los organismos o seres vivos consumen energía para convertir los nutrientes en componentes celulares (anabolismo) y liberan energía al descomponer la materia orgánica (catabolismo).
- Desarrollo. Los organismos aumentan de tamaño al adquirir y procesar los nutrientes. Muchas veces este proceso no se limita a la acumulación de materia sino que implica cambios mayores.
- Reproducción. Es la habilidad de producir copias similares de sí mismos, tanto asexualmente a partir de un único progenitor, como sexualmente a partir de al menos dos progenitores.
- Adaptación. Las especies evolucionan y se adaptan al ambiente.
Autopoiesis
Una forma alternativa de definir a los seres vivos es mediante el concepto de autopoiesis, introducido por los doctores Humberto Maturana y Francisco Varela. La idea es definir a los sistemas vivientes por su organización más que por un conglomerado de funciones. Un sistema se define como autopoiético cuando las moléculas producidas generan la misma red que las produjo y especifican su extensión. Los seres vivos son sistemas que viven mientras conserven su organización. Todos sus cambios estructurales son para adaptarse al medio en el cual ellos existen. Para un observador externo al sistema, esta organización aparece como auto-referida. Las células son los únicos sistemas vivos primarios, es decir aquellos capaces de mantener su autopoiesis en forma autónoma. Los organismos pluricelulares formados por células poseen características similares a las de las células, particularmente el estado estable, pero su vida les es concedida por la organización autopoiética de las células que los constituyen.
Duración de la vida
Uno de los parámetros básicos del organismo es su longevidad. Algunos animales viven tan poco como un día, mientras que algunas plantas pueden vivir millares de años. El envejecimiento puede utilizarse para determinar la edad de la mayoría de los organismos, incluyendo las bacterias.

Desde que en 1665 Robert Hooke describiera la célula como unas cavidades o celdillas en la estructura microscópica del corcho, hubieron de pasar casi dos siglos hasta verificar que todos los organismos animales y vegetales, superiores o inferiores, están formados por células, que son idénticas en lo esencial, pues básicamente constan de los mismos orgánulos y componentes. Este descubrimiento llevó al botánico Schleiden (1838) y al zoólogo Schwann (1839) a la formulación de un principio fundamental de la Biología que se conoce como la teoría celular, cuya enunciación se puede resumir diciendo que la célula es la unidad morfológica y funcional del ser vivo. Sin embargo, la teoría celular no implica necesariamente que todas las células alcancen el mismo nivel de complejidad. Las células más complejas, a las que dedicaremos principalmente nuestra atención en los próximos capítulos, son las células eucariotas o células con núcleo verdadero y la dotación habitual de orgánulos citoplásmicos. Integran organismos tanto una como pluricelulares, a partir de los protozoos. Frente a este mundo de seres vivos, que es el más evidente, existe un mundo microscópico, y a veces submicroscópico, constituido por las células procariotas o células sin verdadero núcleo (aunque sí con ácidos nucleicos) y cuyos orgánulos citoplásmicos quedan limitados a los ribosomas y algunos sistemas simples de membranas. Incluso por debajo de este nivel de organización están los virus y viroides, considerados seres vivos infecciosos, pero no organismos celulares. Existen otros agentes infecciosos que ni siquiera pueden ser considerados seres vivos, como es el caso de los priones, que son proteínas malformadas capaces de contagiar esa malformación, por lo que trataremos de ellos cuando estudiemos la formación de las proteínas.
Virus y viroides
En 1892, Ivanowsky demostró la existencia de unos organismos infecciosos tan pequeños que atravesaban filtros de porcelana con poros menores de un micrómetro. Estos microorganismos son los virus, que ocupan el nivel inferior de organización de los seres vivos. Los virus no son propiamente células. Se ha discutido si realmente son seres vivos, pues dependen de otras células para su reproducción, así como para la síntesis de macromoléculas y otras muchas actividades. No tienen ninguno de los orgánulos que estudiaremos al tratar de la célula y, en cierto sentido, carecen de metabolismo. Para su duplicación dependen de su capacidad de penetrar en células de organismos vivos y de modificar el metabolismo de éstas, reemplazando algunas funciones del DNA celular por las regidas por su propio ácido nucleico. Las funciones de la célula huésped pueden desviarse por completo hacia la producción de nuevos componentes del virus, en cuyo caso el ácido nucleico vírico es el modelo de producción. Se conocen muchos virus diferentes, desde el tamaño de un ribosoma (30 nm) hasta 100-300 nm. Esencialmente constan de un nucleoide envuelto por una cápsula proteica (cápside) que lo protege

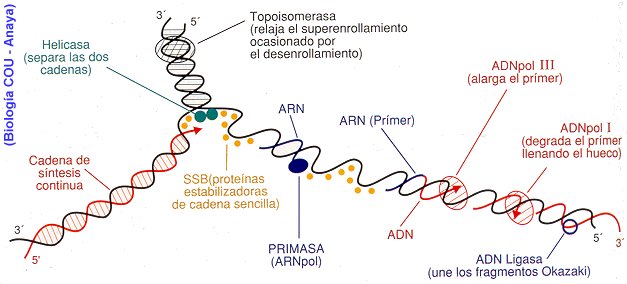
(Fig. 1.15). El nucleoide es un ácido nucleico que puede ser DNA o RNA, pero nunca ambos. Tanto en los virus DNA como en los RNA, el ácido nucleico forma una hélice doble o simple, con disposición lineal o, más rara vez, circular (Tabla 1.2). La capacidad del ácido nucleico vírico es muy variable. Algunos virus, como el del mosaico del tabaco, poseen más de 6000 bases (unos 2 Mm de longitud) en un solo filamento de RNA, que únicamente pueden fabricar tres o cuatro proteínas diferentes. Otros virus, como el bacteriófago T4, llegan a tener hasta 200 000 pares de bases (unos 65 Mm) de doble helicoide de DNA y pueden producir unas 300 proteínas diferentes. La cápsula proteica que envuelve al nucleoide comprende desde 60 hasta miles de moléculas. Los virus más complejos pueden tener, además, una envuelta con fosfolípidos y glucoproteínas recubriendo la cápsula. Las unidades proteicas (capsómeras) muestran generalmente configuraciones geométricas poliédricas. Los componentes de la cápsula son proteínas específicas que sólo se producen bajo la influencia del genoma del virus. Algunos virus tienen una estructura más compleja, como el bacteriófago T4, con cabeza (DNA más envuelta proteica), pieza intermedia, cola y fibras caudales. La cola está provista de una placa terminal con la que el virus se fija a la superficie de la bacteria. En ella hay enzimas que lisan esta superficie para inyectar el ácido nucleico en la bacteria. El resto del bacteriófago queda fuera (Fig. 1.15). Los virus penetran también en la célula por vesiculación (endocitosis). La liberación del virus puede ocurrir al romperse la célula infectada cuando es muy grande el número de virus, o por exocitosis

(Fig. 1.16). La replicación vírica depende del tipo de nucleoide. Los virus DNA (Fig. 1.17) pierden la cápsula al entrar en la célula, donde el DNA se replica originando muchas copias que formarán los nucleoides de los descendientes del virus original. Para formar la cápsula de esos nucleoides, el DNA vírico se transcribe (utilizando las enzimas de la célula) en el RNA mensajero que codifica la síntesis de las proteínas capsulares. Esta síntesis se realiza también utilizando enzimas y ribosomas de la célula. El último paso es el montaje de cada cápsula con su nucleoide y así los nuevos virus quedan finalizados. Algunos virus RNA replican su RNA a expensas de la célula, pero utilizando como modelo su propio RNA vírico, ya que carecen de DNA (Fig. 1.18). Éste es el caso del virus de la gripe y del virus de la estomatitis vesicular, que constituyen un sistema extraordinario en Biología, pues en ningún otro ser se puede sintetizar RNA a partir de RNA. Ello implica la inducción de nuevas enzimas, como las capaces de emplear moldes de RNA. Los ribosomas y otros componentes de la célula se utilizan para las funciones víricas. Lo primero que hace el RNA vírico es actuar como mensajero y emplear los ribosomas para fabricar la enzima RNA replicasa y otras proteínas. Esto permite que el RNA vírico forme múltiples copias de sí mismo, las cuales servirán de modelos para obtener aún más copias de ese RNA. El RNA vírico actúa también como RNA mensajero para la síntesis de las proteínas capsulares que se ensamblarán con las multiples copias obtenidas del RNA.

TABLA 1.2. Características del nucleoide de diferentes virus
Figura 1.15. Representación esquemática de diversos virus. Los tamaños a los que han sido dibujados no guardan proporción entre sí. Virus mosaico del Tabaco, Adenovirus, Virus de inmunodeficiencia humana HIV-1
Otros virus RNA utilizan un mecanismo diferente para su replicación (Fig. 1.19). Son los retrovirus, que parasitan células animales y pueden ser oncogénicos. Poseen la enzima transcriptasa inversa, que también penetra en la célula junto con el nucleoide vírico. Esta enzima permite copiar el RNA vírico, pero no en RNA sino en DNA, el cual se incorpora al DNA de la célula (transducción del DNA) y se transcribe en múltiples copias de RNA, que formarán los nucleoides de los nuevos virus. Además, como en el caso anterior, el RNA vírico actúa como mensajero para la síntesis de las proteínas capsulares.
Hay estructuras constituidas por un ácido nucleico que son aún más simples que los virus. Se trata de los viroides, que consisten en una molécula desnuda de RNA de entre 240 y 600 nucleótidos (de 80 a 200 nm de longitud) e infectan diversas plantas como patatas, cocoteros y crisantemos. No se ha demostrado que este RNA codifique proteína alguna. Cualquier actividad bioquímica en la que esté implicado el viroide requiere las proteínas enzimáticas celulares. Parece que para la propia duplicación del viroide se utiliza la enzima RNA polimerasa II de la célula huésped (que normalmente transcribe el DNA celular en RNA mensajero).
Células procariotas
Son células muy pequeñas, como los micoplasmas, las bacterias típicas como Escherichia coli y las algas verdes azuladas (cianobacterias). Su organización es más sencilla que la de las células eucariotas. Contienen ribosomas y algunos sistemas de membranas que no forman verdaderos orgánulos. En general, el metabolismo de las células procariotas presenta una diversidad extraordinaria. Así como los ciclos metabólicos de algunas bacterias apenas difieren de los de organismos superiores, otras bacterias poseen sistemas metabólicos exclusivos. Esta diversidad explica que se puedan encontrar bacterias en ambientes muy diversos, e indica el alto grado de organización que se esconde en esa aparente sencillez morfológica. La clorofila bacteriana y otros pigmentos fotosintéticos difieren de los de las células eucariotas. Hoy día todos los organismos procariontes se clasifican en dos grandes grupos o subreinos: arqueobacterias o bacterias primitivas, y eubacterias o bacterias verdaderas.
Arqueobacterias
Cumplen con las características generales de las células procariotas al presentar una membrana plasmática (en este caso rodeada de pared celular) que contiene un nucleoide (doble helicoide de DNA no unido a proteínas) y ribosomas de 70 unidades S. No hay orgánulos membranosos. La principal característica de las arqueobacterias es su localización habitual en ambientes que son hostiles para la mayoría de los organismos y donde desarrollan un metabolismo peculiar, de acuerdo con el cual se clasifican en tres grupos:
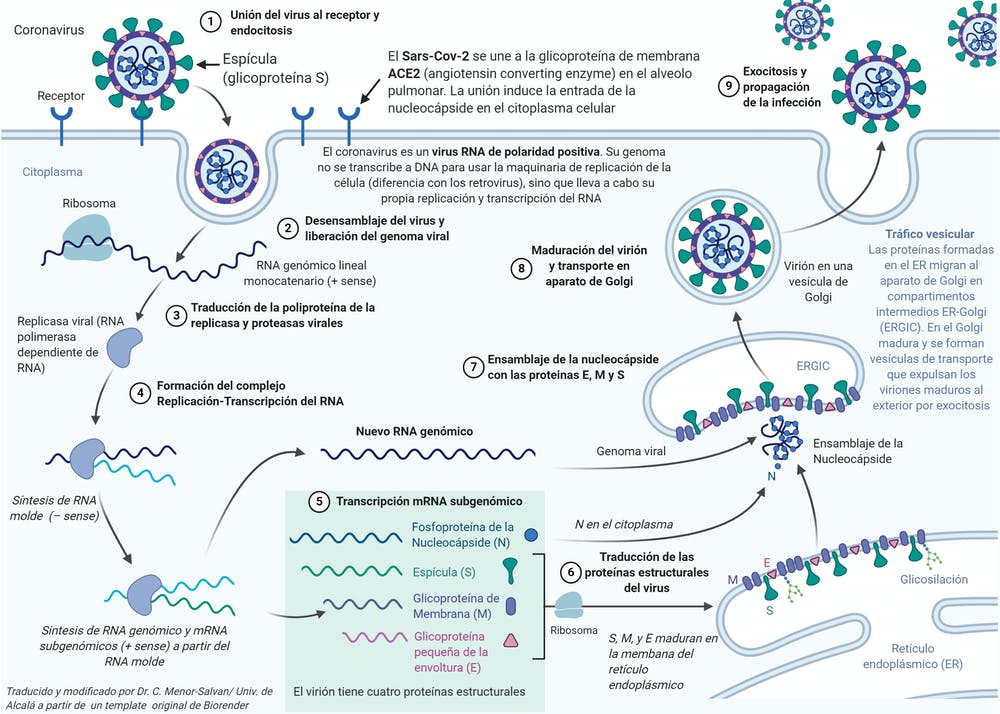
Figura 1.19. Esquema del comportamiento de un virus RNA con un simple helicoide que posee la enzima transcriptasa inversa.
1. Metanógenas. Convierten el CO2 y el H2 en metano (CH4). No pueden vivir en presencia de oxígeno. Se encuentran en las aguas estancadas y residuales (son las que producen el gas de los pantanos), en el fondo del océano y en las aguas termales. También residen en el tracto intestinal de algunos animales, como los rumiantes. 2. Halófitas. Viven en medios salinos, al borde de los océanos y en lagos salados. Algunas crecen incluso en la salmuera saturada. Decoloran y alteran el pescado salado. Mantienen fuertes gradientes de la concentración iónica a través de la membrana plasmática y utilizan esos gradientes para transportar sustancias hacia fuera o hacia dentro de la célula. Algunas halófitas realizan fotosíntesis mediante un pigmento diferente de la clorofila, denominado rodopsina bacteriana por su parecido con el pigmento visual retiniano del mismo nombre.
3. Termoacidófilas. Se encuentran en manantiales calientes y ácidos. Un grupo de estas bacterias (las sulfobacterias) se encuentra en aguas sulfurosas, a unos 80 °C y de pH inferior a 2. Son aerobias y realizan la oxidación de SH2 a S (en vez de la fotosíntesis de H2O a O2). Necesitan esta elevada temperatura para establecer gradientes de electrones que les permitan mantener un pH interno próximo a la neutralidad. Se considera que en estos ambientes debieron de vivir las formas más primitivas de vida antes de que evolucionaran al enriquecerse la atmósfera en oxígeno. Las arqueobacterias no sólo se diferencian de las eubacterias en su especial ambiente y metabolismo, sino también en algunas características moleculares:
1. La pared celular carece de peptidoglucanos, constantes en las eubacterias con pared celular.
2. Los lípidos de la membrana plasmática consisten en un diéter establecido por el glicerol con dos moléculas de fitanol (un alcohol que presenta una cadena alifática ramificada). En las eubacterias y células eucariotas, el glicerol se une por enlaces del tipo éster a cadenas alifáticas rectas.
3. El RNA de transferencia (tRNA) posee una pseudopurina en vez de la base timina presente en las eubacterias y procariotas.
4. La enzima RNA polimerasa difiere de la de las eubacterias.
5. La síntesis proteica se inicia con el aminoácido metionina y no es inhibida por el cloranfenicol. En ambos aspectos las arqueobacterias coinciden con las células eucariotas y se diferencian de las eubacterias.
EUBACTERIAS
Constituyen un grupo muy amplio de microorganismos que comprenden desde las formas más simples, como los micoplasmas, hasta las más complejas, como las cianobacterias, pasando por los organismos que han sido considerados bacterias en sentido estricto, uno de cuyos representantes más estudiado y utilizado en los laboratorios es Escherichia coli. Su metabolismo puede ser autótrofo o heterótrofo. Micoplasmas Pasteur pensó que la pleuroneumonía bovina era causada por un agente bacteriano que no pudo ser aislado por ser demasiado pequeño, de un tamaño comparable al de los virus. Sin embargo, se demostró que este microorganismo no era un virus porque pudo ser aislado y cultivado en un medio sin células. Utilizando filtros de calibre sumamente pequeño, se determinó el tamaño de este microorganismo, que resultó ser de entre 125 y 150 nm, es decir, más pequeño que algunos virus. Se han aislado más de 30 cepas que causan enfermedades respiratorias en aves de corral, artritis en cerdos e infección de ubre en ovejas. En los seres humanos causa pleuroneumonía y uretritis. El género recibió el nombre de Mycoplasma. Son bien conocidas dos especies: M. laidlawii y M. gallisepticum. El M. laidlawii mide de 200 nm a 1 Mm. Tiene vida libre y no necesita células para la duplicación. Contiene DNA, ribosomas y algunas enzimas, como las necesarias para la conversión de la glucosa en ácido pirúvico. Tiene un gránulo de significado incierto y una vacuola que algunos interpretan como pliegue de la membrana plasmática. Todo ello va envuelto en una membrana plasmática de 10 nm de espesor, constituida por una bicapa lipídica que contiene fosfolípidos y colesterol, como las membranas de las células eucariotas
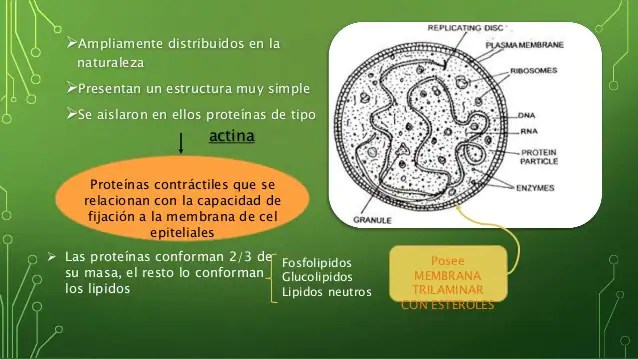
Figura 1.20. Esquema de Mycoplasma laidlawii
Figura 1.21. Imágenes de microscopía electrónica de procariotas. A: Mycoplasma penetrans. No se observan orgánulos celulares sino tan sólo el nucleoide (flecha). X70 000. (Tomado de Yáñez A. y colaboradores. Emerging Infectious Diseases, 1999; 5:1.) B: Escherichia coli. El nucleoide aparece como finas fibrillas (flecha). X40 000. (Micrografía de W. Villiger.) C: Klebsiella oxytoca (bacteria gramnegativa). Se observan la membrana interna (MI), el espacio periplásmico (P), la membrana externa (ME) y la cápsula de polisacáridos (C). X180 000. (Cortesía de A. Martínez y M. Ivarte. Departamento de Histología, Universidad de Navarra.) D: Microscopía electrónica de barrido de Escherichia coli que muestra numerosas fimbrias (flecha). X20 000. (Micrografía de Indigo Instrumens.) E: Micrococo en división. La flecha señala el tabique que separa ambas células hijas. (Micrografía de P. Johson. EMIF. University of Rhode Island.) F: Cianobacteria. L: laminillas. C: cianosomas. X40 000. (Micrografía de Indigo Instrumens.)

(Figs. 1.20 y 1.21.A). Estos microorganismos son los únicos procariotas conocidos que carecen de pared celular. El DNA (4% del peso seco) es un doble helicoide de unos 500 000 pares de bases (unos 170 Mm). El RNA (8% del peso) se encuentra formando parte de algunos centenares de ribosomas de 70 S. Las bacterias típicas tienen 10 veces más bases en su nucleoide y el número de ribosomas es 100 veces mayor. Se calcula que este micoplasma puede sintetizar entre 500 y 1000 proteínas diferentes; entre ellas están las enzimas necesarias para la replicación, transcripción y traducción del DNA, y para el glucólisis anaerobio. En M. laidlawii la reproducción parece ocurrir por formación de nuevas células en su interior que terminarían por liberarse de la célula madre, ya que en el citoplasma de las formas de mayor tamaño se observan inclusiones correspondientes a las formas de menor tamaño. En la especie M. gallisepticum, cuyo tamaño es constante (250 nm), la división celular se produce por tabicación, mediante la formación de un tabique transversal y la escisión posterior. El descubrimiento de estas células planteó esta pregunta: ¿pueden existir células más pequeñas? Tales células podrían revelarse mediante el microscopio electrónico, cultivos o enfermedades. Para ello es necesario que formen agregados de al menos 100 000 moléculas. Al ser la membrana plasmática de unos 10 nm, el diámetro celular debe ser, como mínimo, de 30 nm. Los cálculos bioquímicos aseguran que estos organismos necesitan realizar unas 100 reacciones enzimáticas diferentes. Esto implica un diámetro de unos 40 nm. Además, hay que calcular más moléculas para repuesto de las que se desgastan, lo que nos lleva a un diámetro de unos 50 nm. Pues bien, M. laidlawii tiene el doble de diámetro. Esto sitúa los micoplasmas en el límite de las células vivientes, aunque no hay que descartar la posibilidad de que existan otras células cuyas necesidades bioquímicas sean inferiores a las de las células que conocemos.
Bacterias típicas: Escherichia coli
Una de las bacterias típicas más estudiadas por genetistas y bioquímicos es la Escherichia coli, un bacilo que se encuentra en el tubo digestivo de los mamíferos. Mide 1 s 2 Mm y contiene unos 5000 compuestos diferentes, desde agua a DNA y enzimas (Figs. 1.21.B y 1.22). Se reproduce cada 15-30 minutos en un medio de cultivo compuesto por glucosa y unas sales inorgánicas. Esto quiere decir que posee todas las enzimas necesarias para sintetizar todos sus compuestos a partir de unos pocos elementos. Las bacterias poseen una membrana plasmática de dimensiones normales, recubierta de una pared celular varias veces más gruesa (Figs. 1.21.C, 1.21.D y 1.22). En las bacterias gramnegativas, como E. coli, la pared celular se organiza del modo siguiente: externamente a la membrana plasmática o membrana interna, hay una capa denominada espacio periplástico, constituida por proteínas y peptidoglucanos; estos últimos son polisacáridos que forman cadenas lineales (generalmente de dos tipos de azúcares repetidos), unidas entre sí por cortos péptidos. Más externamente se encuentra una nueva bicapa lipídica (la membrana externa) que contiene una proteína transmembranosa llamada porina, que hace que la membrana sea permeable a muchas sustancias. Recubriendo la membrana externa puede haber un revestimiento desarrollado de hidratos de carbono, denominado cápsula, constituido por glucolípidos, esto es, glúcidos unidos a los fosfolípidos de la hemimembrana lipídica externa de esta membrana externa (Figs. 1.21.C, 1.22 y 1.23). En los potentes antígenos que presenta la membrana externa radica fundamentalmente la toxicidad que muestran las bacterias gramnegativas en los mamíferos, así como su mayor resistencia a la penicilina y a la enzima lisozima, que impiden la agregación de los componentes de la pared. A esta estructura de la pared celular se debe el que la tinción gram sea negativa en estos organismos porque, después de teñirlos con el colorante cristal violeta, son decolorados por disolventes debido a que éstos pueden atravesar la pared celular. La pared celular de las bacterias grampositivas es más gruesa, y consta de una única bicapa lipídica (no hay membrana externa), recubierta de una gruesa capa de peptidoglucanos que no pueden atravesar los disolventes. El DNA se encuentra mucho más centralizado que en los micoplasmas. Es una doble cadena de unos 3 millo nes de pares de bases (1000 a 1500 Mm de longitud). En E. coli, en fases próximas a la división celular, el DNA forma dos regiones nucleares, cada una en contacto con una invaginación llena de repliegues de la membrana plasmática llamada mesosoma, que es el esbozo del primer orgánulo celular. Las bacterias como E. coli pueden sintetizar miles de proteínas diferentes y poseer hasta 10 000 ribosomas. Éstos son de 70 S y contienen mayor proporción de RNA que de proteínas. El mesosoma interviene en la duplicación y distribución del DNA y parece que contiene enzimas relacionadas con este proceso. De hecho, se duplica con el DNA, previamente a la división celular. Además, el mesosoma contiene las enzimas de la cadena transportadora de electrones (en los micoplasmas estarían adosadas a la membrana plasmática). Otras muchas enzimas se encuentran en el citoplasma o adosadas a la membrana plasmática. No hay fagocitosis ni pinocitosis. Tampoco existen lisosomas. Sin embargo, algunas bacterias producen exoenzimas, que actúan fuera de la membrana plasmática desdoblando macromoléculas en otras más sencillas que puedan ser incorporadas a través de la membrana. También poseen enzimas que destruyen tejidos facilitando la penetración bacteriana. Sin embargo, la existencia de estas enzimas no supone la unión de vesículas a la membrana plasmática. La presencia de numerosos ribosomas adheridos a ésta sugiere que estas enzimas se sintetizarían en la misma membrana y se verterían fuera de la célula. Muchas bacterias poseen flagelos, que son muy diferentes de los de las células eucariotas.

Figura 1.22. Representación esquemática de Escherichia coli.
Consisten en una fibra única (triple hélice), de 10 a 20 nm de espesor y varios micrómetros de largo, que nace en un gránulo basal. La fibra está formada por la proteína flagelina. La rotación de los flagelos ayuda al desplazamiento de la bacteria. Otros apéndices son fibrillas sumamente finas y cortas denominadas fimbrias o pili (Fig. 1.21.D). Las bacterias se dividen por división directa. Una vez replicado el DNA, las dos copias se separan por estrangulación de la membrana entre los puntos de anclaje de una y otra copia, formándose dos células hijas, cada una con un nucleoide idéntico (Fig. 1.21.E). Algunas bacterias pueden transformarse en esporas inactivas, extraordinariamente resistentes a grandes cambios de temperatura, deshidratación, etcétera. Las esporas tienen una región nuclear, poco citoplasma y una pared muy gruesa, que se forma a expensas de la membrana plasmática y la pared celular. En la germinación funcionan ciertos mecanismos de síntesis antes inactivos.
Figura 1.23. Pared celular de bacteria gramnegativa.
Cianobacterias, Aunque la mayoría de las algas son eucariotas, las llamadas algas verde azuladas (cianobacterias) son procariotas. Se encuentran como células independientes o como colonias pluricelulares filamentosas. Comparten con otras bacterias muchas de sus características como la presencia de DNA de doble helicoide y de ribosomas libres de 70 S (Figs. 1.21.F y 1.24). La pared celular es semejante a la bacteriana, en concreto a la de las bacterias gramnegativas. Una característica propia, ausente en otras bacterias y que revela un nivel de organización discretamente superior, es la existencia de un metabolismo fotosintético semejante al de las plantas verdes. Además de la clorofila, contienen otro pigmento, la ficobilina, que comprende, a su vez, dos pigmentos: ficocianina (azul) y ficoeritrina (rojo). Separando fracciones por centrifugación, se ha comprobado que la clorofila está dentro de unos sáculos membranosos (laminillas), mientras que la ficobilina se encuentra en gránulos denominados cianosomas adosados a las membranas. Cianosomas y laminillas pueden realizar la fotosíntesis juntos, pero no por separado. Estos pigmentos permiten que estas algas puedan desarrollarse en condiciones ambientales extremas de temperatura y concentración salina. Las membranas con clorofila, sin que puedan considerarse verdaderos cloroplastos, constituyen un orgánulo membranoso de células procariotas. Además, las cianobacterias pueden fijar N2 convirtiéndolo en NH3, con el que sintetizan sustancias orgánicas nitrogenadas como aminoácidos o nucleótidos. Pueden vivir prácticamente con sólo luz, N2, CO2 y agua.

| 4 | Membrana exterior Membrana plasmática Nucleoide Vacuolas Tilacoides Ficobilisomas Pared de peptidoglucanos Citoplasma | 🙁 |
| 8 | Membrana exterior Membrana plasmática Nucleoide Vacuolas Tilacoides Ficobilisomas Pared de peptidoglucanos Citoplasma | 🙁 |
| 3 | Membrana plasmática | 🙂 |
| 5 | Membrana exterior Membrana plasmática Nucleoide Vacuolas Tilacoides Ficobilisomas Pared de peptidoglucanos Citoplasma | 🙁 |
| 1 | Membrana exterior | 🙂 |
| 2 | Pared de peptidoglucanos | 🙂 |
| 6 | Membrana exterior Membrana plasmática Nucleoide Vacuolas Tilacoides Ficobilisomas Pared de peptidoglucanos Citoplasma | 🙁 |
| 7 | Vacuolas |
Figura 1.24. Representación de una Cianobacteria.
CÉLULAS EUCARIOTAS En las páginas anteriores se han resumido los diferentes niveles de organización en la estructura de los seres vivos. En los virus sólo hay ácido nucleico y cápsula proteica. No hay ribosomas ni síntesis proteica; tampoco vida libre. Los restantes microorganismos mencionados son procariotas (pro = previo, carios = núcleo), con un núcleo no organizado, y todos poseen una membrana plasmática que alberga un contenido: DNA, ribosomas (de 70 S), proteínas y enzimas. En las bacterias típicas existe, además, una pared celular y un esbozo de orgánulo: el mesosoma. Las cianobacterias aportan a este nivel de organización la presencia de membranas dentro de la célula, las cuales podrían considerarse como el primer orgánulo. Las células eucariotas suelen ser hasta 1000 veces más voluminosas que las procariotas

(Fig. 1.25). El DNA es también unas 1000 veces más abundante, va unido a unas proteínas denominadas histonas y está encerrado en una doble membrana. Ésta constituye una envoltura nuclear completa que forma parte del sistema de membranas interno de la célula, con el que mantiene conexión en las células recién formadas (Fig. 1.26). En el núcleo se encuentra el nucléolo que es la expresión morfológica de la síntesis de ribosomas. El sistema de membranas celulares alcanza un notable desarrollo y es en él donde se realizan gran parte de las reacciones vitales. Todas las sustancias que la célula necesita penetran a través de la membrana plasmática; algunos de los materiales lo hacen en el interior de vesículas de endocitosis, que se invaginan desde la membrana plasmática (Fig. 1.26). En las células animales, que necesitan aumentar su superficie (para el transporte de sustancias) sin que se produzca paralelamente un aumento del volumen celular, la membrana plasmática desarrolla pliegues y proyecciones digitiformes de ésta (microvellosidades) (Fig. 1.27.A). En las células vegetales, cuya membrana plasmática se adapta a la pared celular rígida, el aumento de la superficie celular se consigue mediante el desarrollo de una gran vacuola intracitoplásmica (Fig. 1.27.B). El sistema de membranas internas, además de formar la envoltura nuclear, constituye una compleja red de canales por el citoplasma denominada retículo endoplasmático. Los ribosomas son de 80 S y, en contraste con los de las células procariotas, poseen más proteínas que RNA. Estos ribosomas sintetizan proteínas, que quedan en el citoplasma fundamental (también denominado hialoplasma o citosol), y se adosan al retículo endoplasmático (y a veces a la membrana nuclear externa) formando el retículo endoplasmático rugoso, en cuyo caso, las proteínas sintetizadas se almacenan en el interior de estas membranas y parte de ellas serán exportadas fuera de la célula. Otra variedad de retículo endoplasmático es el retículo endoplasmático liso, que carece de ribosomas y, que, entre otras funciones, es el lugar de síntesis de muchos lípidos y sus derivados. El retículo endoplasmático conecta con otras membranas que forman grupos de sáculos superpuestos que constituyen el aparato o complejo de Golgi, donde se producen modificaciones en las moléculas fabricadas en el retículo endoplasmático, principalmente en los hidratos de carbono añadidos a las proteínas. Del complejo de Golgi emigran vesículas de exocitosis hacia la membrana plasmática, a la que se fusionan vertiendo hacia el exterior parte de las sustancias fabricadas por la célula y que constituyen secreciones celulares. Del complejo de Golgi emanan también vesículas que contienen las enzimas necesarias para la digestión intracelular y que se denominan lisosomas. Éstos forman compartimientos aislados de los restantes componentes celulares, que no pueden así ser alcanzados por las enzimas digestivas. Otras vesículas con una función muy diferente son los peroxisomas, donde se generan y degradan peróxidos peligrosamente reactivos durante la oxidación por el oxígeno de diversas moléculas. Orgánulos membranosos, aparentemente no conectados con los anteriores y que poseen cierta autonomía debida a un DNA y RNA propios, son los cloroplastos, que realizan la fotosíntesis (se encuentran sólo en las células vegetales con esta capacidad) y las mitocondrias, que realizan el metabolismo oxidativo.

Figura 1.25. Electronografías de células eucariotas. A: Un organismo unicelular: la microalga Dunaliella bioculata. CP: cloroplasto; F: flagelo; G: complejo de Golgi; GA: gránulo de almidón; MT: mitocondria. N: núcleo; P: pirenoide; PG: plasto glóbulo; RER: retículo endoplasmático rugoso; R: Ribosomas; V: vacuola. X10 500. (Micrografía de K.A. Bérubé. Tomada de European Microscopy and Analysis 1993; 26: 21.) B: Células de organismo superior (ganglio linfático humano). Se observan el núcleo (N), mitocondrias (M), cisternas aisladas de retículo endoplasmático rugoso (RER), ribosomas libres (R), un complejo de Golgi (G) y una pareja de centriolos (C). X5000.
Finamente, las células eucariotas poseen un citoesqueleto, constituido por microtúbulos, microfilamentos y filamentos intermedios. Estas estructuras permiten a la célula mantener su forma, y poder cambiarla y desplazarse, así como para mantener la ubicación de los orgánulos celulares o facilitar el desplazamiento de éstos y de otros componentes y sustancias. La motilidad celular, o del medio en contacto con la célula, se consigue mediante cilios y flagelos (Fig. 1.27.A). En la Tabla 1.3 se expresan las principales diferencias entre las células eucariotas y las procariotas.

Figura 1.26. Representación esquemática tridimensional de una célula eucariota.
Figura 1.27. Esquema de células eucariotas vistas con el microscopio electrónico. A: Célula animal (epitelial). B: Célula vegetal (parénquima clorofílico).
La transición de células procariotas a eucariotas no es brusca. En los protozoos diplomonados y microsporidios se encuentran los modelos más sencillos de célula eucariota. Quizá el más simple de todos sea el diplomonado Giardia, un organismo unicelular que vive de forma anaeróbica como parásito intestinal de mamíferos. Posee núcleo verdadero (de hecho, es binucleada) y citoesqueleto, pero carece de retículo endoplasmático, complejo de Golgi, mitocondrias y cloroplastos.

TABLA 1.3. Diferencias entre células procariotas y eucariotas
ESTRUCTURA DE LAS MEMBRANAS CELULARES
La membrana plasmática no es visible con el microscopio de luz, que sólo permite apreciar las sustancias que la rodean; por eso, su estudio comenzó con métodos indirectos. En 1895, Overton presumió la existencia de una membrana de naturaleza lipídica en la célula debido a que la superficie celular es fácilmente traspasada por lípidos y muy resistente al paso de corriente eléctrica. En 1897, Langmuir estudió el comportamiento de los fosfolípidos al extenderlos sobre el agua, y observó que los grupos polares (hidrófilos) de cada molécula quedaban en contacto con la superficie acuosa mientras que los grupos no polares (hidrófobos) se disponían perpendicularmente a ésta. Si se añadía otra capa de fosfolípidos, ésta se disponía enfrentada a la anterior para que los grupos polares y no polares quedasen también en la misma relación respecto al agua. En 1925, Gorter y Grendel extrajeron los lípidos de la membrana de eritrocitos y calcularon que, al extenderlos sobre el agua, ocupaban una superficie doble de la que debían ocupar las membranas de los eritrocitos. Llegaron a la conclusión de que la membrana es una capa lipídica bimolecular (Fig. 2.1.A). En 1932, Cole estudió la tensión superficial de membranas de huevos de erizo de mar. El valor encontrado resultó inferior a la tensión superficial teórica para una capa de fosfolípidos; de ello dedujo que éstos deberían ir acompañados de proteínas que disminuyeran su tensión superficial. En 1935, Danielli y Dawson propusieron un modelo de estructura de la membrana plasmática en el que las proteínas se sitúan con los grupos polares de la bicapa lipídica. Posteriormente, incluyeron en su modelo poros o canales en la membrana para explicar el paso de sustancias (Fig. 2.1.B).

Figura 2.1. Primeros modelos de membrana plasmática. A: Bicapa lipídica que forma la estructura básica de la membrana plasmática. Los grupos polares (hidrófilos) se disponen en la periferia, mientras que los grupos no polares (hidrófobos) se disponen en el interior de la membrana. B: Modelo de membrana plasmática propuesto por Danielli y Dawson. Las proteínas rodean a los lípidos. C-D: Resumen de otros modelos de membrana en los que las proteínas no rodean a los lípidos, sino que están embebidos en ellos. Los lípidos se disponen formando esferas (C) o en bicapa (D). El modelo C se aplicó principalmente a membranas citoplásmicas; el D, a la plasmática. Del modelo C podría pasarse con facilidad al D en los procesos de unión de vesículas a la membrana plasmática.
ESTUDIOS CON EL MICROSCOPIO ELECTRÓNICO
Con la aplicación del microscopio electrónico al estudio de las células se vio por primera vez la membrana plasmática. Ésta, observada a grandes aumentos (por encima de los 40 000), aparece como una estructura trilaminar. Inicialmente, el espesor de la membrana plasmática se estimó en 7.5 nm. Al mejorar la calidad de la fijación se corrigió este valor situándolo en 10 nm, valor admitido actualmente y corroborado por diversos métodos de estudio. La misma imagen trilaminar de la membrana plasmática se observaba también en las membranas de los orgánulos citoplásmicos (membranas citoplásmicas), por lo que Robertson (1959) denominó a esta estructura unidad de membrana (Fig. 2.2), al entender que existe una igualdad esencial entre todas las membranas celulares. En 1962, Stoeckenius elaboró una membrana artificial con dos capas de fosfolípidos, la fijó con osmio y la observó con el microscopio electrónico. La imagen resultante era la estructura trilaminar, por lo que se llegó a la conclusión de que la fijación del osmio tenía lugar en los grupos polares de los fosfolípidos. Al añadir proteínas a esta membrana, se repetía la imagen anterior pero con las líneas densas más gruesas, por lo que se pensó que las proteínas se disponían junto con los grupos polares de los fosfolípidos. La idea de que todas las membranas celulares son esencialmente iguales se compagina con la observación de una continuidad entre diferentes membranas celulares: entre el retículo endoplasmático liso y el rugoso; entre éste y el complejo de Golgi; entre las vesículas que surgen del complejo de Golgi y la membrana plasmática; y entre ésta y las vesículas que se adentran en el citoplasma. También se observan conexiones entre el retículo endoplasmático rugoso y la envoltura nuclear, la cual, en su membrana externa, tiene ribosomas adheridos. Incluso en células jóvenes y, con un carácter temporal, se ha visto una continuidad entre el retículo endoplasmático rugoso y la membrana plasmática. Sin embargo, era lógico pensar también en componentes singulares de acuerdo con la especialización del orgánulo. De hecho, nunca se han visto conexiones entre mitocondrias o cloroplastos y otras membranas. Un hecho significativo es que las membranas citoplásmicas son más delgadas que la membrana plasmática: su espesor es de unos 7 nm, frente a los 10 nm de la membrana plasmática. El espesor de la bicapa lipídica es también menor: 4 nm en las membranas citoplásmicas y 5 nm en la membrana plasmática. Además, las membranas citoplásmicas tienen, en general, mayor proporción de proteínas que la plasmática. Esto último es muy evidente en las membranas mitocondriales que conservan su estructura al eliminar los lípidos. Por otra parte, esta estructura trilaminar no está tan clara en algunas membranas citoplásmicas, particularmente en el caso de las mitocondrias, cuya membrana muestra partículas globulares, sobre todo cuando se utilizan determinados fijadores. Esto llevó a algunos investigadores de la década de 1960-70 a proponer diferentes modelos, en los que los lípidos de la membrana se disponían radialmente formando glóbulos, con los grupos polares hacia el exterior (Fig. 2.1.C). Al mismo tiempo, en estos modelos se discutía la disposición de las proteínas, que se distribuirían por toda la membrana (Figs. 2.1.C. y 2.1.D).
El mosaico fluido de membrana. estructura y composición
Hacia 1970 se produjeron grandes avances en la búsqueda de un modelo de membrana debido al desarrollo del concepto termodinámico de interacciones hidrófobas e hidrófilas entre moléculas, enlaces no covalentes como puentes de hidrógeno, e interacciones electrostáticas. Se llegó a la conclusión de que las proteínas (que también tienen grupos polares y no polares) no podían disponerse en configuración beta, como se postulaba en el modelo de Danielli y Dawson, sino de modo que, como ocurre en los lípidos, sus grupos polares estén en contacto con la fase acuosa y los no polares queden en el interior de la membrana. Por lo que respecta a las técnicas de microscopía electrónica, las dos siguientes fueron de un gran valor en el estudio de las membranas:
1. Contraste negativo, que permitió observar protuberancias e irregularidades en las membranas, imposibles de apreciar en los cortes. En algunas células se vio que la membrana plasmática está constituida por bloques o unidades poligonales.
2. Criofractura-réplica. Al romperse las membranas por las líneas de mínima resistencia, éstas quedan divididas en dos hemimembranas:
- P (protoplásmica o interna)
- E (exoplásmica o externa).
La superficie interna de cada hemimembrana no es lisa, y sobre ella resaltan partículas de 4 a 16 nm sobre un fondo liso. Estas partículas se corresponden con cavidades en el fragmento complementario de la membrana y son debidas a proteínas, por lo que son más abundantes en membranas ricas en enzimas.
Figura 2.2. A-B: Membranas plasmáticas y membranas citoplásmicas que muestran la estructura trilaminar de ambas. Flechas grandes: hemimembranas externas de la membrana plasmática. Flechas pequeñas: hemimembranas internas de la membrana plasmática. Estrella: espacio intercelular. En el citoplasma (C) de ambas células se observan algunas membranas citoplásmicas de retículo endoplasmático liso (cabezas de flecha) y de mitocondrias (flechas abiertas). A: X125 000. B: X200 000.
Las dos partes de la membrana son asimétricas, siendo las partículas más abundantes y mayores en la hemimembrana P (Fig. 2.3).

Figura 2.3. Muestra obtenida por criofractura-réplica de la membrana plasmática. El plano de fractura divide a ésta en dos hemimembranas que presentan una estructura asimétrica. La hemimembrana señalada como I (P) es la interna o protoplásmica; la señalada como E es la externa o exoplásmica. Sobre la hemimembrana interna se observan partículas de mayor tamaño que las que aparecen sobre la externa, la cual presenta cavidades en las que encajan las partículas que sobresalen en la hemimembrana protoplásmica. X90 000. (Tomado de Fawcett DW. A Textbook of Histology, 11.ª ed. Philadelphia, Saunders, 1986.)
Con los resultados de estos y otros estudios, Singer y Nicolson (1972) llegaron a proponer un modelo que ha sustituido a todos los anteriores y que, pese al tiempo transcurrido, se encuentra en vigor y se aplica a todas las membranas de la célula. Es el modelo del mosaico fluido de membrana (Figs. 2.4 y 2.5), en el que las proteínas, lípidos e hidratos de carbono se sitúan en una configuración estable de baja energía libre. Los lípidos forman una bicapa en la que se disponen las proteínas configuradas de acuerdo con las interacciones que establecen con las moléculas que las rodean. Hay también oligosacáridos que se disponen sobre los lípidos y las proteínas en la hemimembrana E. La membrana plasmática del eritrocito de rata tiene un 60% de proteínas y un 40% de lípidos. Esta proporción es similar a la encontrada en las membranas plasmáticas de la mayoría de los tipos celulares; por ejemplo, la membrana plasmática del hepatocito de rata tiene un 58% de proteínas y un 42% de lípidos. No obstante, existen membranas plasmáticas que se alejan mucho de esta proporción, como la mielina, que tiene un 20% de proteínas y un 80% de lípidos. Las membranas citoplásmicas, además de ser más delgadas que la plasmática, como ya se ha dicho, poseen mayor proporción proteínas/lípidos. La diferencia más notable radica en la membrana mitocondrial interna, que tiene un 80% de proteínas y un 20% de lípidos. La Tabla 2.1 muestra la composición de diferentes membranas celulares.
Lípidos de las membranas
Los lípidos forman una doble capa, con los grupos hidrófobos en el centro y los hidrófilos en el exterior, en contacto con la fase acuosa. Forman la matriz de la membrana. Hay unos cinco millones de moléculas lipídicas por Mm2 de membrana. Los principales tipos de lípidos que forman ambas bicapas de la membrana son los siguientes (Fig. 2.6):
1. Las grasas neutras, formadas por ésteres de glicerol y uno (monoglicéridos), dos (diglicéridos) o tres (triglicéridos) ácidos grasos, son un componente minoritario, o incluso ausente, en la membrana plasmática. En las membranas de bacterias y células vegetales son frecuentes los glucolípidos simples, constituidos por glicerol esterificado con uno o dos ácidos grasos y con un monosacárido o un oligosacárido unido al tercer hidroxilo.
2. Fosfolípidos. Son fosfodiglicéridos, es decir, consisten en una molécula de glicerol esterificado con dos ácidos grasos (de 16 y 20 carbonos de longitud); cada uno de ellos se encuentra unido por su extremo carboxilo a un hidroxilo del glicerol. El tercer hidroxilo del glicerol está sterificado con un fosfato. Si este fosfato no se esterifica con ningún otro componente, el fosfolípido se denomina ácido fosfatídico. Si el fosfato se une a otros radicales, se denomina de acuerdo con este radical. Los principales fosfolípidos son los unidos a colina (fosfatidil colina o lecitina), serina (fosfatidil serina o cefalina), etanolamina (fosfatidil etanolamina, también denominada cefalina, como el fosfolípido anterior), inositol (fosfatidil inositol), o a otra molécula de glicerol (fosfatidil glicerol). En las membranas también hay otro fosfolípido, formado por la unión de dos ácidos fosfatídicos a otra molécula de glicerol (difosfatidil glicerol o cardiolipina).
3. Esfingolípidos. Son derivados de la esfingosina, que es un aminodialcohol con un largo hidrocar
Figura 2.4. Representación tridimensional del modelo del mosaico fluido de membrana. La membrana se ha separado parcialmente en dos hemimembranas, como queda tras el tratamiento de criofractura-réplica.
Figura 2.5. Modelo de membrana plasmática basado en el modelo del mosaico fluido propuesto por Singer y Nicolson. Las proteínas integrales aparecen embebidas en la bicapa lipídica formando hélices de uno o varios pasos. La mayoría de ellas atraviesan totalmente la membrana (proteínas transmembranosas); otras sólo ocupan una hemimembrana. Las proteínas periféricas están adosadas a una hemimembrana, uniéndose a una proteína integral, a un lípido o a un oligosacárido que, a su vez, está unido a un lípido. Los oligosacáridos quedan sobre la hemimembrana externa y forman el glicocálix; pueden estar unidos a una proteína integral o a un lípido
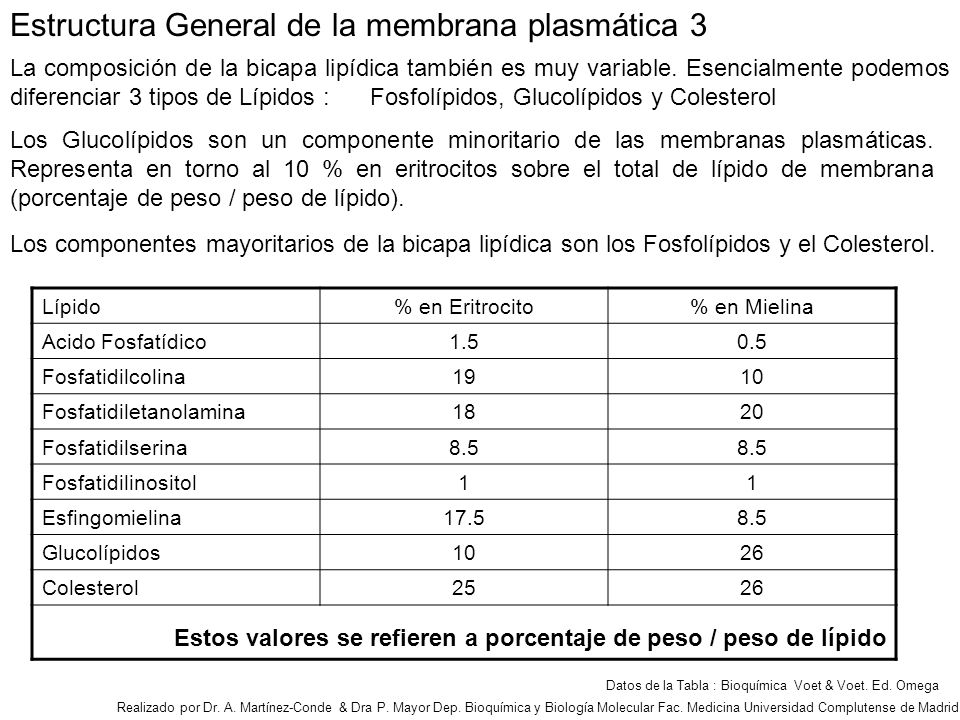
TABLA 2.1. Diferencias en la composición de lípidos (%) entre diferentes tipos de membranas celulares en el hígado de rata.
Figura 2.6. Representación esquemática de la composición de los lípidos de la membrana plasmática.
buro terminal. La esfingosina unida a un ácido graso en su grupo amino forma la ceramida.
— La ceramida esterificada con fosfato y colina en el grupo hidroxilo terminal forma la esfingomielina. Debido a la presencia del fosfato y colina, se puede incluir entre los fosfolípidos.
— La ceramida unida a hidratos de carbono (de uno a 15 azúcares) forma glucolípidos complejos (glucoesfingolípidos), abundantes en las membranas de células animales.
Éstos pueden ser:
• Cerebrósidos. El hidrato de carbono es un monosacárido. En la mielina abunda el galactocerebrósido, que sólo tiene galactosa.
• Gangliósidos. El hidrato de carbono es un oligosacárido con varios residuos de ácido siálico (N-acetil neuramínico), que le confiere carga negativa.
4. Esteroles derivados del ciclopentano-perhidro-fenantreno, con un hidroxilo en un extremo y una cadena alifática corta en el otro. El más común es el colesterol. Entre los diferentes modelos experimentales de membrana utilizados para investigar las propiedades de la bicapa lipídica se encuentran los liposomas, constituidos por bicapas lipídicas en forma de esfera de 25 nm a 1 Mm de diámetro, y las membranas negras, que son bicapas lipídicas planas formadas en un agujero situado en la separación entre dos compartimientos acuosos. Estos estudios han llevado a precisar que las membranas funcionales requieren una matriz lipídica fluida; esto es, una membrana es funcional cuando se mantiene por encima del punto de fusión de sus lípidos. La temperatura de fusión depende de la longitud de la cadena de los fosfolípidos, del número de dobles enlaces y de la concentración de colesterol. En esta matriz fluida, los lípidos pueden hacer desplazamientos de difusión lateral, rotación y flexión con una constante de difusión lateral de 10–8 cm2 /s. En contraste con estos movimientos laterales, son infrecuentes los movimientos de voltereta (flip-flop), esto es, inversión de la polaridad de las moléculas cruzando la membrana de arriba a abajo. Sólo son frecuentes durante la síntesis de membrana por el retículo endoplasmático. La permeabilidad de la membrana disminuye con la abundancia de colesterol, que presenta los siguientes efectos:
(a) dificulta que las cadenas de hidrocarburos de los ácidos grasos se junten y cristalicen;
(b) dificulta la permeabilidad de la membrana para las pequeñas moléculas solubles; y
(c) aumenta la flexibilidad y la estabilidad mecánica de la bicapa.
Existe asimetría en la bicapa lipídica, pues hay mayor proporción de fosfatidil colina y esfingomielina (fosfolípidos con colina y que poseen ácidos grasos saturados) en la hemimembrana exoplásmica (E), y mayor cantidad de fosfatidil etanolamina y fosfatidil serina (fosfolípidos con ácidos grasos insaturados) en la hemimembrana citoplásmica (P). La matriz lipídica de la hemimembrana P es más fluida que la de la E debido a su mayor contenido en ácidos grasos insaturados. La mayor presencia de fosfatidil serina (con fuerte carga negativa) en la hemimembrana P determina que exista una diferencia de carga entre ambas hemimembranas.
Proteínas de las membranas
Proteínas integrales y periféricas
De acuerdo con configuración que adoptan en la membrana, las proteínas son de dos tipos (véase Fig. 2.5): Proteínas integrales
Estas proteínas suelen atravesar por completo la membrana (proteínas transmembranosas) formando hélices A de paso único o múltiple por la membrana, aunque se conocen algunas que sólo ocupan una hemimembrana, como el citocromo b5 del retículo endoplasmático. Son anfipáticas, es decir, presentan una distribución asimétrica de los grupos hidrófilos e hidrófobos, lo que las capacita para estar en parte embebidas y en parte sobresalientes en la membrana. La extensión en que una proteína integral está embebida se regula por el equilibrio termodinámico; esto es, está determinada por la secuencia de aminoácidos y la estructura covalente de la molécula, así como por su interacción con las moléculas que la rodean. Los grupos polares de la proteína quedan generalmente en la superficie de la membrana, mientras que los residuos no polares permanecen embebidos entre las cadenas hidrocarbonadas de los fosfolípidos. Las proteínas integrales están firmemente unidas a los lípidos por interacciones hidrófobas. Algunas refuerzan la unión por enlaces covalentes con lípidos y sólo se disocian de éstos por tratamientos drásticos (detergentes, agentes que desnaturalizan las proteínas y disolventes orgánicos) que destruyen la integridad de las membranas. Las proteínas integrales pueden realizar movimientos de rotación y de traslación con una constante de difusión de aproximadamente 5 s 10–9 cm2/s. El desplazamiento de las proteínas se mostró con el siguiente experimento. Se cultivaron conjuntamente células de ratón y células humanas y se marcó una proteína específica de la membrana plasmática de la célula de ratón mediante un anticuerpo unido a un colorante fluorescente. Se hizo lo mismo con otra proteína específica de la membrana de la célula humana, pero marcando ésta con un colorante diferente para distinguir ambas proteínas. Tras fusionar ambas células, se vio que, al principio, cada tipo de proteína quedaba limitada a un área diferente en la célula híbrida, pero posteriormente ambas proteínas se entremezclaban (Fig. 2.7). Durante estos movimientos las proteínas son capaces de mantener su orientación molecular y su grado de intercalación en la membrana, como resultado de su estructura anfipática.
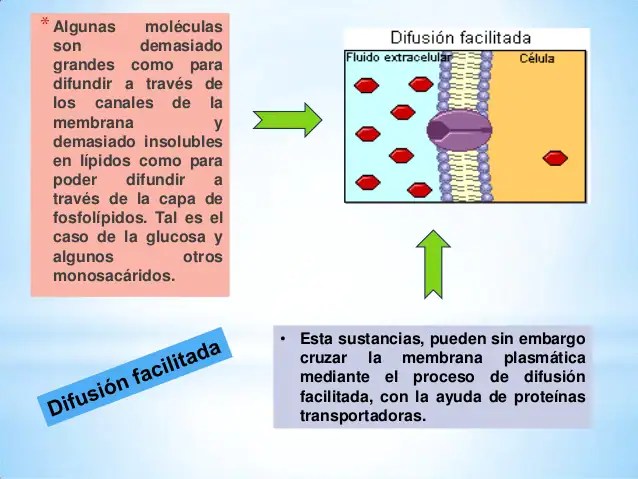
Figura 2.7. Demostración del desplazamiento de las proteínas por la membrana plasmática. Cada una de las dos células en cultivo presenta una proteína diferente marcada, utilizando dos colorantes diferentes para que se distingan. Al fusionar ambas células, en la célula híbrida resultante se observan las dos proteínas, al principio en áreas diferentes; después, al mantener un tiempo el cultivo a 37°C, ambas proteínas se entremezclan.
El movimiento de desplazamiento de las proteínas puede abrir poros transitorios o dinámicos. De esta manera, el paso a través de la membrana del agua y de sustancias insolubles en lípidos no requiere la presencia de poros, que nunca se han visto. Además, algunas proteínas actúan como moléculas portadoras uniéndose a las sustancias transportadas. Son proteínas transmembranosas que poseen actividades tipo permeasa, y en su interior existen caminos polares para permitir el transporte molecular. Proteínas periféricas Estas proteínas no son transmembranosas y sobresalen en una de ambas hemimembranas (véase Fig. 2.5). Están asociadas a la membrana únicamente por un enlace covalente con un ácido graso o por interacciones no covalentes (principalmente electrostáticas) con una proteína integral. No están estrechamente asociadas a los lípidos, y un tratamiento con la adición de un agente quelante (tratamiento de tipo suave), basta para disociarlas enteras de la membrana. Se encuentran sobre todo en la hemimembrana P y corresponden en su mayor parte a enzimas. En la hemimembrana E son muy escasas y pueden unirse a una molécula de GPI (glucosilfosfatidil inositol), esto es, a un oligosacárido que, a su vez, se encuentra unido a los dos ácidos grasos de una molécula de fosfatidil inositol. Algunas proteínas específicas de la membrana plasmática Las proteínas de la membrana plasmática más estudiadas son las del eritrocito, debido a la facilidad con que pueden obtenerse: basta con provocar la lisis de esta célula, en un medio hipotónico. Estas proteínas comprenden (Fig. 2.8):
1. La glucoforina. Es una glucoproteína de unos 30 kilodaltons (kDa), cuya porción transmembranosa tiene la forma de una hélice A de paso único. El segmento externo está muy glucosilado, con oligosacáridos que están unidos a oxígeno o a nitrógeno y constituyen hasta un 64% del peso de la proteína. Estos oligosacáridos constituyen casi la totalidad de los hidratos de carbono de la membrana del eritrocito; predomina (hasta el 90%) el ácido siálico, que tiene fuerte carga negativa.
2. La proteína banda
3. Es también transmembranosa y está formada por dos monómeros de 100 kDa (929 aminoácidos) cada uno, que forman múltiples hélices A transmembrana (unos 14 segmentos) y proporcionan un canal hidrófilo para el paso de Cl– y CO3H–. En su cara externa presenta algunos oligosacáridos unidos a nitrógeno. Se dispone formando tetrámeros (dos dímeros) y es crucial en el paso del CO2 de la sangre al interior del eritrocito.
3. La anquirina o proteína banda 2.1, de 210 kDa. Conecta cada dímero banda 3 a una proteína esquelética llamada espectrina.
4. Las proteínas esqueléticas espectrina, actina y tropomiosina, situadas bajo la membrana plasmática, y las proteínas banda 4.1 y aducina, que se encuentran asociadas a las anteriores. La espectrina supone el 25% de las proteínas de esta membrana. Es una molécula larga, delgada y flexible, constituida por dos monómeros alargados (A de 240 kDa y B de 220 kDa) que se disponen formando una doble hélice de unos 100 nm de longitud y unos 5 nm de espesor. A su vez, dos dímeros se asocian en sentido longitudinal y opuesto, estableciendo contacto por sus cabezas, para formar tetrámeros. A este contacto se ancla la espectrina en la membrana mediante la proteína anquirina, la cual, como se ha dicho, se ancla a su vez en la dímera banda 3. Los tetrámeros de espectrina forman una red bajo la membrana plasmática asociándose con sus colas. A esas colas se adosan filamentos cortos de actina (35 nm de longitud y 6 nm de grosor), unidos a la proteína tropomiosina (43 kDa) y a tres moléculas de la proteína banda 4.1 (82 kDa). Finalmente, a cada banda 4.1 se une una molécula de aducina. De la actina y tropomiosina, así como de otras proteínas que forman parte del citoesqueleto de las células eucariotas.
5. Una proteína periférica interna: la enzima gliceraldehído 3 fosfato deshidrogenasa. La membrana plasmática de la mayoría de las células nucleadas de eucariotas contiene otras proteínas distintas cuya organización difiere en cada caso. Algunas de esas proteínas sirven de receptores que intervienen en procesos de reconocimiento y adhesión celular; otras actúan como transportadores hacia el interior o el exterior de la célula; otras son enzimas que catalizan reacciones asociadas con la membrana y, finalmente, otras son proteínas estructurales que, junto con los receptores, conectan la membrana plasmática con el citoesqueleto, con otra célula adyacente o con la matriz extracelular. En membranas plasmáticas de diversos tipos celulares se han detectado cerca de 30 enzimas. Las más constantes son: 5’ nucleotidasa, Mg2+ATPasa, Mg2+ ATPasa activada por Na+ y K+, fosfatasa alcalina, adenilato ciclasa, fosfomonoesterasa ácida y RNAasa. L
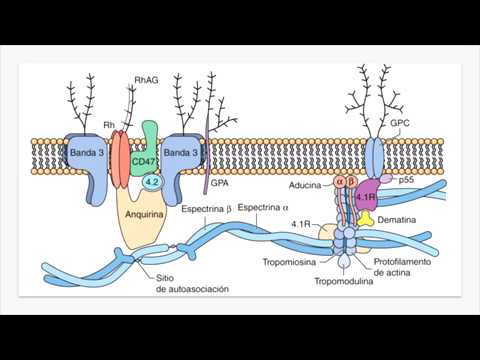
Figura 2.8. Representación de las proteínas esqueléticas asociadas a la membrana plasmática del eritrocito de mamífero.
plasmática carece de cadena respiratoria y tampoco posee actividad glucolítica. Determinadas enzimas de la membrana plasmática están limitadas a la superficie donde desempeñan su función (apical, lateral o basal). Las uniones estrechas entre células impiden el desplazamiento de estas proteínas. La composición lipídica en cada una de esas zonas de membrana puede ser también diferente. El citoesqueleto asociado a la membrana plasmática también difiere del encontrado en el eritrocito, y consiste generalmente en filamentos de actina, de mayor longitud que los del eritrocito. Estos filamentos se entrecruzan formando una red que se une a la membrana de diversas formas. En estas uniones intervienen proteínas especializadas. En las membranas citoplásmicas las proteínas varían dependiendo del orgánulo membranoso. De ellas se tratará al estudiar cada uno de ellos.
Hidratos de carbono de las membranas (glicocálix)
Estructura y composición Los hidratos de carbono están presentes en la membrana plasmática unidos covalentemente a proteínas (glucoproteínas) o a lípidos (glucolípidos). Se encuentran del lado externo y son generalmente oligosacáridos. La célula queda así recubierta por una envoltura de material hidrocarbonado, denominado glicocálix (véanse Figs. 2.4 y 2.5), que es particularmente visible en algunas células (véase Fig. 2.25.B) y que llega a representar entre el 2 y el 10% del peso de la membrana. En esta cubierta también pueden encontrarse algunas proteínas. Las más conocidas son las ya mencionadas, que se unen al GPI y actúan como receptores de señales extracelulares. También puede haber glucoproteínas y proteoglucanos (algunos unidos también al fosfatidil inositol) que fueron segregados por la célula al espacio extracelular y luego adsorbidos por la superficie celular. La diferencia entre las glucoproteínas y los proteoglucanos radica en que, mientras que las glucoproteínas constan de un polipéptido unido a uno o escasos oligosacáridos, los proteoglucanos presentan numerosas y largas cadenas hidrocarbonadas (disacáridos repetidos muchas veces) unidos a la cadena polipeptídica (véase Fig. 7.5). Casi todas las proteínas presentan oligosacáridos en su lado externo, pero sólo la décima parte de las moléculas lipídicas de la hemimembrana externa están unidas a oligosacáridos. Como en la membrana hay unos 50 veces más lípidos que proteínas, habría que pensar que hay cinco veces más oligosacáridos unidos a lípidos que a proteínas. Sin embargo, el peso total de los oligosacáridos unidos a proteínas es mayor que el de los unidos a lípidos, pues mientras que cada molécula de lípido portadora de oligosacáridos posee una única cadena de hidrato de carbono, cada molécula de proteína posee varias cadenas. Los oligosacáridos del glicocálix unidos a proteínas se unen bien al nitrógeno (en un residuo del aminoácido asparagina, por lo que se denominan oligosacáridos N), o al oxígeno (en un residuo del aminoácido treonina o serina, por lo que se llaman oligosacáridos O). Los primeros presentan 12 azúcares y son ricos en manosa, mientras que los segundos sólo tienen unos cuatro azúcares. Una forma de identificar los hidratos de carbono del glicocálix son las proteínas denominadas lectinas, de la familia de las selectinas, que se unen específicamente a secuencias determinadas de hidratos de carbono. Los oligosacáridos del glicocálix unidos a lípidos forman principalmente glucoesfingolípidos, pero también hay oligosacáridos unidos a fosfatidil inositol. El grado de desarrollo del glicocálix es muy variable. En la mayoría de las células el glicocálix forma una delicada capa, difícilmente apreciable con el microscopio electrónico, pero en algunas células epiteliales está muy desarrollado. Uno de los ejemplos mejor estudiados es el glicocálix del epitelio intestinal. Estas células poseen numerosas prolongaciones citoplásmicas hacia la luz, denominadas microvellosidades, que están recubiertas por un material filamentoso, el cual, visto con el microscopio electrónico de gran poder de resolución, se presenta como unos filamentos muy ramificados formando un entramado tridimensional. Se le denomina glicocálix antenular (Fig. 2.25.B). Funciones, Aunque todas las células poseen glicocálix, éste no es igualmente visible en todas ellas ni responde a las mismas necesidades. Las principales funciones reconocidas en el glicocálix son las siguientes:
1. Es responsable de la carga negativa de la superficie celular, principalmente debida al ácidosiá y de los cambios en la carga eléctrica del medio extracelular, actuando como una resina intercambiadora de iones.
2. Reconocimiento y fijación de las partículas que incorpora la célula por endocitosis.
3. Reconocimiento específico de células entre sí durante el desarrollo embrionario, permitiendo la agrupación de las células para generar los tejidos y órganos. La implantación de la metástasis depende de la capacidad de las células tumorales no sólo para emigrar sino también para crecer en lugares nuevos, rodeadas de células con las que normalmente no interaccionan. De esta manera, estas células deben ignorar las propiedades del glicocálix en el reconocimiento y la diferenciación.
4. Participación en las uniones de células entre sí y con la matriz extracelular efectuadas por glucoproteínas transmembranosas como cadherinas (unen células entre sí) e integrinas (unen células a la matriz extracelular).
5. Propiedades inmunológicas. Contiene muchos de los antígenos celulares que causan el rechazo de trasplantes e injertos. Un ejemplo son los grupos sanguíneos que residen en el glicocálix de los eritrocitos.
6. Anclaje de enzimas. En el glicocálix de algunas células hay unidades globulares (5-6 nm de diámetro) que contienen enzimas, como leucoaminopeptidasas en los hepatocitos y maltasa en los enterocitos (células del epitelio intestinal).
Balsas lipídicas de la membrana plasmática
Los esfingolípidos, que tienen largas cadenas de ácidos grasos saturados, crean intensas fuerzas de van der Waals que atraen los lípidos adyacentes formándose microdominios transitorios, ricos en esfingolípidos y colesterol, de unos 70 nm de diámetro, también denominados balsas lipídicas (Fig. 2.9). Esas zonas ocupan ambas hemimembranas, son algo más gruesas que el resto de la bicapa, concentran componentes del glicocálix y acomodan determinadas proteínas. Algunas de éstas son largas proteínas transmembranosas que se unen a oligosacáridos que, a su vez, se unen a las ya mencionadas lectinas. Hay también proteínas transmembranosas que sirven para la transmisión de señales extracelulares al citosol, proteínas periféricas de la hemimembrana externa que se unen a un GPI, y proteínas del medio extracelular que se concentran para ser incorporadas a la célula.
Renovación de las membranas celulares
Marcando las proteínas con leucina-3 H se ha visto que los polipéptidos de alto peso molecular de la membrana plasmática se renuevan cada 2-5 días, mientras que los de bajo peso molecular lo hacen cada 7-13 días. Mar cando radiactivamente los lípidos se prueba que éstos se renuevan cada 3-5 días. La membrana plasmática se encuentra en un continuo proceso de reciclaje. De ella se invaginan vesículas con contenidos necesarios para el metabolismo de las células (endocitosis), lo que supone una pérdida de membrana, y a ella se fusionan vesículas procedentes del citoplasma (principalmente del complejo de Golgi) (exocitosis), lo que supone una recuperación de membrana (Fig. 2.10). La renovación de la membrana plasmática a partir de vesículas del complejo de Golgi exige, a su vez, un incremento de las membranas de este orgánulo para reponer las membranas perdidas. Estas nuevas membranas proceden del retículo endoplasmático, que es el lugar de síntesis de las membranas celulares (con excepción de las membranas de las mitocondrias, de los plastidios y quizá de los peroxisomas). Por otra parte, las membranas de las vesículas de endocitosis terminan uniéndose a lisosomas que, a su vez, reciben membranas del complejo de Golgi (cargadas con enzimas lisosómicas) y emiten membranas hacia éste mediante vesículas con los receptores para cargar enzimas lisosómicas en el complejo de Golgi.
Síntesis de las membranas celulares
Como la formación de las membranas requiere no sólo lípidos sino también proteínas, la síntesis de los com ponentes de las membranas citoplásmicas se realiza en el retículo endoplasmático liso y rugoso: liso en cuanto que posee enzimas para sintetizar fosfolípidos; rugoso, porque debe poseer ribosomas para sintetizar las proteínas integrales. Las proteínas periféricas internas se sintetizan en ribosomas libres (no en el retículo endoplasmático rugoso) próximos a la membrana plasmática. La glucosilación de las proteínas cuyos hidratos de carbono formarán parte del glicocálix se inicia en el retículo endoplasmático rugoso y se completa en el complejo de Golgi. En éste también se produce la glucosilación de los lípidos, completando el glicocálix. Los fosfolípidos y el colesterol, los dos elementos constitutivos principales de todas las membranas celulares, se sintetizan en el retículo endoplasmático liso a partir de los ácidos grasos formados en el hialoplasma. Las moléculas lipídicas recién sintetizadas se sitúan en la hemimembrana P (del lado del hialoplasma). La translocación de la mitad de estos lípidos a la hemimembrana E tiene lugar mediante una translocasa de fosfolípidos denominada escramblasa, que también se encuentran en la membrana plasmática y que equilibra ambas hemimembranas en pocos minutos (Fig. 2.11). La escramblasa cataliza el movimiento flip-flop de fosfatidil colina, fosfatidil serina y fosfatidil inositol, pero no el de fosfatidil etanolamina. Algunos de los fosfolípidos se transforman en lípidos con etanolamina una vez translocados, pero esto ocurre en pequeña proporción, de modo que se establece una asimetría en la composición lipídica de la membrana del retículo endo plasmático y de todas las membranas derivadas de éste, incluida la membrana plasmática. En esta última, además de la escramblasa, hay otra proteína, denominada flipasa, que es exclusiva de la membrana plasmática y mueve fosfatidil serina y fosfatidil etanolamina desde la hemimembrana E hacia la P, contribuyendo a la asimetría. En el retículo endoplasmático liso se forma también la esfingosina por la condensación de serina y un ácido graso. La esfingosina unida a otro ácido graso forma la ceramida (véase Fig. 2.6). A partir de la membrana del retículo endoplasmático se forman las membranas del complejo de Golgi. En la hemimembrana E de éste, se forman la esfingomielina (si a la ceramida se añade fosfato y colina tomados de la fosfatidil colina) y los glucoesfingolípidos (si a la ceramida se añaden monosacáridos para formar cerebrósidos u oligosacáridos para formar gangliósidos) (véanse Figs. 2.6 y 2.11). Como en el complejo de Golgi no hay translocasas de fosfolípidos, tanto la esfingomielina como los glucolípidos permane cen en la hemimembrana E donde fueron formados. También en el complejo de Golgi se produce la glucosilación del fosfatidil inositol de la hemimembrana E.

Figura 2.9. Representación de un dominio o balsa lipídica de la membrana plasmática. Estas zonas tienen un espesor mayor que el del resto de la membrana y son ricas en esfingolípidos y colesterol. Presentan un glicocálix muy denso y largas proteínas transmembranosas unidas a oligosacáridos con gran afinidad por las lectinas. Hay también proteínas periféricas externas que se unen al glucosilfosfatidil inositol.

Figura 2.10. Renovación de la membrana plasmática a partir de las vesículas emanadas del complejo de Golgi. Nótese cómo los oligosacáridos que se encuentran en el interior de las vesículas pasan a situarse en la superficie de la membrana plasmática formando el glicocálix al producirse la inversión de la curvatura de la membrana.
Con excepción de las mitocondrias y cloroplastos, que como se verá más adelante son orgánulos semiautónomos, y quizá también de los peroxisomas, los demás sistemas de membranas de la célula se hallan interconectados, bien directamente o a través de vesículas que transportan membrana y sustancias de un sistema a otro. Cuando los fosfolípidos van a formar parte de la membrana de las mitocondrias hace falta un procedimiento especial de transferencia (véase página 210). En el caso de los cloroplastos no se plantea este problema, pues fabrican sus propios lípidos. Por lo que respecta a las proteínas, tanto la mitocondria como los cloroplastos son capaces de sintetizar algunas de ellas con sus propios ribosomas, pero la mayoría deben ser sintetizadas en el citosol. De la transferencia de proteínas a mitocondrias y cloroplastos se tratará también en las páginas 208 y 217.

Figura 2.11. Esquema de la formación de la bicapa lipídica en la membrana del retículo endoplasmático. A partir de los ácidos grasos (sintetizados en el hialoplasma) se originan los fosfolípidos, que se disponen formando una monocapa, continuación de la hemimembrana P del retículo endoplasmático. Posteriormente, una translocasa de fosfolípidos (escramblasa) pasa parte de los nuevos fosfolípidos con colina, serina o inositol a la otra hemimembrana. Los fosfolípidos con etanolamina no pueden ser translocados, pero algunos de los anteriores pueden transformarse en fosfatidil etanolamina. Algunas moléculas de fosfatidil serina se unen a un ácido graso y forman esfingosina que, con otro ácido graso, da lugar a ceramida. Una vez que la membrana ha pasado al complejo de Golgi, la ceramida puede convertirse en esfingomielina (por unión a fosfato y colina) o en glucoesfingolípidos (por unión a monosacáridos y oligosacáridos). Ambos tipos de moléculas se forman en la hemimembrana E del complejo de Golgi, donde permanecen definitivamente pues este orgánulo carece de translocasas.
Permeabilidad de la membrana plasmática. difusión simple
Las membranas celulares se comportan como membranas semipermeables; es decir, el agua se mueve con mayor facilidad que la mayoría de los solutos y se desplaza hacia donde éstos están más concentrados. Este proceso se llama ósmosis. El agua tiende a entrar en las células, donde la concentración de iones y pequeñas moléculas es mayor que en el medio externo. Para compensar esa entrada de agua, las células han desarrollado diferentes estrategias, como la presencia de paredes celulares rígidas (bacterias, células vegetales), de orgánulos activos en la expulsión de agua (vacuolas pulsátiles) o de bombas de membrana. Por otra parte, además del agua, muchas otras moléculas pueden atravesar la membrana plasmática. La velocidad de penetración de una molécula a través de la membrana plasmática (permeabilidad) varía ampliamente entre las diferentes moléculas. Una molécula atraviesa más rápidamente la membrana cuanto menor es su tamaño (menor peso molecular) y mayor es su solubilidad en lípidos con relación a su solubilidad en agua (coeficiente de partición). La membrana plasmática deja pasar con facilidad moléculas pequeñas no polares (oxígeno, nitrógeno, benceno) y moléculas pequeñas polares sin carga (agua, urea, glicerol, CO2); sin embargo, es mucho más impermeable a los iones y moléculas cargadas, por lo que estas sustancias atraviesan la membrana muy lentamente (Fig. 2.12.A). Otras membranas de la célula, así como las bicapas lipídicas artificiales, poseen las mismas propiedades. Aunque el movimiento de estas moléculas se realiza en ambas direcciones, el flujo neto de ellas se produce a favor de gradiente de concentración, aumentando linealmente con el valor del gradiente, lo que se denomina difusión simple.
Transporte mediado por proteínas
La mayoría de las sustancias necesarias para las células (azúcares, aminoácidos, nucleósidos, vitaminas, etc.) son moléculas polares o con carga neta y atraviesan demasiado lentamente las membranas por difusión simple para satisfacer las necesidades de las células. Por ello las células han desarrollado numerosos sistemas de transporte basados en proteínas transmembranosas de paso múltiple (Fig. 2.12.B). Si las moléculas se transportan a favor del gradiente, este proceso ocurre espontáneamente y se denomina transporte pasivo; en cambio, si lo hacen en contra del gradiente el proceso necesita un aporte de energía para poder realizarse, y se habla entonces de transporte activo.

Figura 2.12. Representación del paso de sustancias a través de la membrana plasmática.
A: La membrana plasmática deja pasar con facilidad y a favor del gradiente de concentración, por difusión simple, pequeñas moléculas no polares y pequeñas moléculas polares sin carga; en cambio, es poco permeable a las grandes moléculas polares sin carga y a todas las moléculas cargadas.
B: Para aumentar la velocidad de penetración a través de la membrana (permeabilidad), muchas moléculas polares, como iones, aminoácidos, azúcares y nucleótidos, utilizan sistemas de transporte. En el esquema se representan algunos mecanismos de transporte pasivo que favorecen la difusión simple y permiten la difusión facilitada.
Transporte pasivo
El transporte pasivo puede realizarse bien mediante canales (proteínas de canal) o mediante transportadores (proteínas transportadoras o permeasas) (Fig. 2.12.B).
Proteínas de canal
Las proteínas de canal forman canales acuosos que permiten el paso de moléculas polares o de iones a velocidades muy superiores a las que permitiría su difusión simple a través de la bicapa lipídica. Aunque en algunos canales el flujo de sustancias transportadas aumenta linealmente con el gradiente, siguiendo las leyes de la difusión simple, en la mayoría de ellos, el proceso tiende a saturarse con altas concentraciones, lo que indica que la molécula transportada interactúa con el canal proteico. La mayoría de los canales para el paso de iones actúan como puertas transitorias y su apertura y cierre están regulados por diferentes tipos de estímulos, que pueden ser:
1. La unión a un ligando (una molécula específica del canal y diferente de las sustancias transportadas por éste); son los canales regulados por ligando (Fig. 2.13.A).
2. Un cambio de potencial de membrana; son los canales regulados por voltaje (Fig. 2.13.B).
3. La unión a un nucleótido cíclico, principalmente cGMP; son los canales regulados por nucleótidos cíclicos.
4. El cambio en la concentración de algún ion; son los canales regulados por concentración iónica.
5. La estimulación mecánica. Un ejemplo de la actuación combinada de estos canales es la transmisión sináptica (véase Fig. 7.39).
Proteínas transportadoras.
Difusión facilitada Las proteínas transportadoras, o permeasas, permiten el paso altamente selectivo de determinadas moléculas o iones. Presentan además una cinética de transporte muy diferente de la difusión simple, saturándose el transporte con determinadas concentraciones, por lo que se denomina difusión facilitada (la cinética del transporte por proteínas de canal sería intermedia entre ésta y la de la difusión simple).
Figura 2.13. Dos sistemas de canales que aumentan la permeabilidad en la difusión simple.
A: Proteína de canal regulada por ligando. La unión de éste a la proteína determina la apertura del canal.
B: Proteína de canal regulada por cambios en la polaridad eléctrica de la membrana.
En la difusión facilitada, cuando la proteína transportadora tiene ocupados todos los centros de unión al sustrato se dice que está saturada, y la velocidad de transporte con esa concentración es máxima (Vmáx). Cada transportador tiene una velocidad máxima característica, así como una constante de unión para su soluto (KM), que es igual a la concentración del soluto cuando la velocidad de transporte es la mitad de la máxima (Fig. 2.14). La cinética del transporte mediado por transportadores se parece a la de las reacciones enzimáticas, aunque aquí no se produce modificación del sustrato, sino facilitación del transporte. Muy probablemente, en este proceso se producen también cambios de conformación del transportador. Los transportadores pueden transportar un solo tipo de moléculas (transporte sencillo o uniporte) o simultáneamente dos tipos de moléculas (cotransporte) (Fig. 2.15.A) mediante cambios en la configuración de la molécula transportadora (Fig. 2.15.B). En este segundo caso, la célula puede utilizar el movimiento a favor del gradiente de una de las sustancias transportadas para transportar la otra en contra de su gradiente de concentración, de modo que el transporte global sea energéticamente favorable, como estudiaremos en este mismo capítulo al tratar del transporte activo (véase página 56). Si ambas sustancias se transportan en la misma dirección, el transporte se denomina paralelo o simporte, y si se transportan en direcciones opuestas se habla de transporte antiparalelo o antiporte. Como modelos de la difusión facilitada se han estudiado con detenimiento pequeñas moléculas hidrófobas sintetizadas por microorganismos y utilizadas probablemente como armas biológicas, denominadas ionóforos. Se conocen dos clases de ionóforos: los móviles y los formadores de canales. Ambos actúan protegiendo la carga del ion transportado, de modo que éste pueda penetrar en el interior hidrófobo de la bicapa. Al no estar acoplados a fuentes energéticas, sólo permiten el movimiento neto de los iones a favor del gradiente electroquímico (véase Fig. 2.12.B) Figura 2.15. Proteínas transportadoras en la difusión facilitada. A: El transporte puede ser de una sola molécula (uniporte) o de dos diferentes (cotransporte). En este caso, el transporte puede ser uni o bidireccional. B: Posible mecanismo por el que se realiza el transporte mediante un cambio en la configuración de la proteína transportadora.



{kind=link}