PRIMARIA. SECUNDARIA.
Estructura primaria del tallo en dicotiledóneas
Estructura primaria del tallo en dicotiledóneas. La estructura primaria es la que presentan los entrenudos del tallo mientras crecen en longitud. En esta estructura sólo existen tejidos primarios desarrollados a partir de los meristemas apicales (promeristemos),
Tejidos meristemáticos
Son tejidos embrionarios capaces de diferenciarse o perpetuarse; es decir, se multiplican activamente para formar los tejidos adultos diferenciados (crecimiento y especialización) y a su vez originan nuevas células meristemáticas. Los meristemas permiten que se produzca el crecimiento de las plantas en sentido longitudinal y diametral. El crecimiento longitudinal, también llamado crecimiento primario, se produce por la acción del meristema apical; mientras que el crecimiento diametral o en grosor, también denominado crecimiento secundario, se produce por divisiones que ocurren en el cambium vascular y, en menor proporción, en el cambium cortical.
Clasificación:
Según la posición topográfica en la planta:
Apicales: localizados en el ápice de los órganos (tallos, raíces, glándulas, etc.) que forman.
Basales: localizados en la base de los órganos (p.e., espinas) que forman.
Intercalares: situados entre células diferenciadas (maduras), contribuyendo al crecimiento en ambos lados.
Laterales: localizados en la periferia de los órganos, favoreciendo el crecimiento en grosor.
Axilares: son los meristemas apicales de las yemas, situados en las axilas de las hojas.
Según la naturaleza de las células que lo forman:
Primarios: provienen directamente de células que nunca han perdido su capacidad de división. (derivados de células embrionarias).
Secundarios: se originan a partir de células diferenciadas que nuevamente adquieren su capacidad de división.
Según el tiempo de aparición (secuencia de formación) en la planta:
Primarios: son los meristemas apicales del tallo y raíz presentes originariamente en el embrión y son capaces de producir los tejidos primarios.
Secundarios: se originan a partir de tejidos primarios por desdiferenciación. Producen los tejidos secundarios.
Características de las Células Meristemáticas.
Son células Indiferenciadas, pequeñas e isodiamétricas (excepto cámbium vascular). forman tejidos compactos, sin espacios intercelulares, gran núcleo (difuso) y poco citoplasma, pared celular delgada constituida de pared primaria y lamela media. (Algunos poseen campos de poros los), no poseen inclusiones citoplásmicas y con pocos orgánulos: abundantes ribosomas libres y dictiosomas, retículo endoplásmico (liso y rugoso) escaso. mitocondrias escasas y con pocas crestas, presentan proplastidios.
Meristemas Primarios (facilitan el crecimiento en longitud). Tipos.
Cuando ya la planta está formada, el tejido meristemático sólo se encuentra localizado en determinadas regiones del vegetal llamadas “zonas de crecimiento”, que conservan indefinidamente su carácter embrionario y su capacidad de división. Estas zonas se localizan en las yemas del tallo y en la región subterminal de las raíces. Los meristemos que aparecen en el extremo de los brotes o yemas y permiten el crecimiento en longitud de la planta, reciben el nombre de meristemos apicales.
Entre éstos se encuentran la protodermis, el procámbium, el meristema fundamental o básico.
Meristemas apicales: situados en los ápices de brotes y raíces (tanto principales como laterales), aquí se incluyen los meristemas caulinares, encargados del crecimiento del tallo, ramas y hojas y los meristemas apicales radicales, que son los responsables del crecimiento de las raíces.
Meristemas intercalares: situados en la base de los entrenudos de las ramas. (Aparecen más tarde en el tiempo).
Meristemas apicales caulinares:
Meristemas apicales radicales (yemas radicales):
Periciclo: Meristemo remanente propio de la raíz que se sitúa en la periferia del cilindro vascular. Colabora en la formación del córtex secundario, incluida la peridermis, del cambium vascular, y de las raíces laterales.
Meristemas Secundarios. Tipos. Se hacen cargo de facilitar el crecimiento en grosor.
El cambium vascular: Es un meristemo lateral formado por una monocapa cilíndrica de células situado en aquellos tallos y raíces que van a sufrir engrosamiento secundario. Sus células son de dos tipos: iniciales fusiformes e iniciales radiales, y se dividen en un plano tangencial al tallo o raíz. Las fusiformes dan lugar hacia el interior a células que se diferencian como xilema II y hacia el exterior a células que se diferencian como floema II. Las radiales dan lugar a parénquima. El cambium vascular del tallo se origina de dos formas: Bien a partir de células de procámbium presentes entre la xilema I y el floema I, dando lugar al cambium fascicular, bien por desdiferenciación de células parenquimáticas de los radios medulares, dando lugar al cambium interfascicular.
El cambium fascicular: es el cambium vascular que se forma a partir del procámbium situado entre la xilema y el floema primarios de los haces vasculares.
El cambium interfascicular: Es el cambium vascular que se forma por desdiferenciación de células del parénquima radiomedular, es decir, del parénquima situado entre los haces vasculares.
El cámbium suberógeno o felógeno: Meristemo lateral presente en los tallos y raíces de las plantas con engrosamiento secundario. Es una monocapa cilíndrica de células formada por la desdiferenciación de una capa de parénquima cortical subepidérmico. Por división tangencial estas células generan hacia el interior la felodermis y hacia el exterior el súber (corcho).
Las partes fundamentales que integran a esta estructura son: epidermis, corteza y estela o médula.
La epidermis, es el tejido más externo constituido por una capa delgada de células, se encuentra recubierto por una cutícula impermeable de cutina y, en la mayoría de los casos, pelos o tricomas.
Protodermis
Es una capa de células meristemáticas externa de los ápices vegetativo y radical que dará lugar a la epidermis.
La protodermis es una capa que se desarrolla sobre la superficie del ápice y sólo se divide en sentido anticlinal, es decir, perpendicular a la superficie. A través de la división de este meristema primario se origina la epidermis, la cual cumple las funciones de protección, contra agentes químicos y físicos externos y contra la desecación, además de permitir el intercambio gaseoso, secreción de compuestos metabólicos y absorción de agua en el cuerpo primario de la planta.
Meristema fundamental o básico
Meristema fundamental es un grupo de células meristemáticas que origina el resto de los tejidos de las plantas (parénquimas, colénquimas, esclerénquimas, etc).
Procámbium
El Procámbium meristema primario constituido por una capa de células que originará los tejidos vasculares primarios (xilema y floema primarios). Puede formar, en algunos casos, el cámbium vascular.
Tejidos de protección. Epidermis
La epidermis procede de la capa más externa del promeristemo, la protodermis. Recubre externamente el cuerpo de la planta a modo de envoltura protectora, pero permite al mismo tiempo, el intercambio de materia con el medio exterior. Generalmente consta de una sola capa de células vivas, las que, en visión frontal, presentan aspecto poligonal o alargado y se disponen unidas unas a otras sin dejar espacios intercelulares constituyendo una película continua. A menudo el contorno lateral de las células es ondulado o dentado lo que refuerza la trabazón marginal de los elementos celulares.
La principal función de la epidermis es la de protección contra desecación y ataque de agentes externos, la misma tiene modificaciones como: los pelos o tricomas y estomas.
Tejidos de Protección. Modificaciones de la epidermis. Tricomas
Se da el nombre de tricoma, a todo apéndice epidérmico, independientemente de la forma que este tenga, que se encuentra a modo de resalto en la superficie de los órganos vegetales. Su forma, estructura y funciones, son diversos.
Las formas más comunes de tricomas son los pelos; también se consideran como tales a las papilas, las escamas y pelos absorbentes de las raíces. En cuanto a su estructura pueden ser unicelulares o pluricelulares, sencillos o ramificados y desempeñan diversas funciones: algunos son glandulares y producen sustancias volátiles que actúan como atractivos de insectos polinizadores o como repelentes de otros insectos, otros son urticantes (células de defensa) ya sea porque producen sustancias urticantes (hortiga) o porque tienen una constitución rígida y puntiaguda. Pueden encontrarse en todas las partes de la planta. Los tricomas deben estar formados exclusivamente por células epidérmicas
La corteza o córtex, está formada por todos los tejidos comprendidos entre la epidermis y el cilindro central, estela o médula, de la siguiente forma: la colénquima, ubicado inmediatamente después de la epidermis alternando con parénquima clorofílico;
Tejidos mecánicos o de sostén. Colénquima
Son los tejidos que le dan resistencia y elasticidad a la mayoría de las plantas.
Sin una cierta solidez la planta no podría conservar su forma.
Están constituidos por células dispuestas en una masa compacta y con las paredes celulares fuertemente engrosadas, bien en parte, como en la colénquima, o bien en su totalidad como en el esclerénquima.
Los principales tejidos mecánicos son: la colénquima y el esclerénquima.
La colénquima se caracteriza por ser un tejido vivo con protoplasto poco activo, sus células vistas en una sección longitudinal presentan una forma rectangular y transversalmente el aspecto más poligonal, la pared primaria es más gruesa en las esquinas (constituida por celulosa y sustancias pépticas), raras veces presentan cloroplastos, puede impregnarse de lignina y transformarse en esclerénquima.
La estructura y disposición de las células colenquimáticas en el cuerpo de la planta indican que la función primaria de este tejido es de sostén. Es el tejido típico de sostén, primero de los órganos en crecimiento y segundo de los órganos adultos herbáceos, presentándose característicamente en posición periférica en los tallos y hojas.
La estructura de la membrana o pared celular es el carácter más distintivo de las células colenquimáticas. Los espesamientos se disponen desigualmente, con cierta variabilidad en los distintos tipos de plantas. Una forma más común es presentar los espesamientos más importantes en los ángulos (colénquima angular), donde se reúnen varias células. Si el espesamiento no es tan manifiesto en los ángulos la cavidad celular adquiere una forma circular en su sección transversal (colénquima laminar). Son también conocidos como angular, laminar y lagunar.
El colénquima más especializado se origina conjuntamente con los tejidos vasculares a partir del procámbium y el menos especializado a partir del meristemo fundamental.
Tejidos parenquimáticos. Clorofílico
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas, presenta de las funciones metabólicas vacuola grande.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
El parénquima clorofílico es el parénquima que realiza una de las funciones más importantes del vegetal, la fijación del carbono utilizando las radiaciones solares. Ocupa las partes su epidérmicas de la planta hasta donde pueda penetrar la luz, posee células siempre vivas provistas de cloroplastos. Se encuentra más desarrollado en las hojas, donde forma una capa de tejido en empalizada y de parénquima lagunar.
Plastidios, Cloroplastos
Además del núcleo y las vacuolas, los cloroplastos, constituyen los organelos más conspicuos de una célula vegetal. Los cloroplastos están rodeados por una doble membrana, con una estructura interna constituida por un sistema de membranas, separadas por una matriz de naturaleza proteica llamada estroma, poseen DNA con una estructura similar al encontrado en células procarióticas, así como ribosomas, embebidos en el estroma. Todos los plastidios se desarrollan a partir de pro cloroplastos, que son cuerpos pequeños encontrados en plantas que crecen tanto en la luz como en la oscuridad. Se dividen por fisión o bipartición, como lo hacen las mitocondrias y los organismos procariotas. Los plastidios incoloros se conocen como leucoplastos, contienen enzimas responsables de la síntesis del almidón. Los leucoplastos mejor conocidos son los amiloplastos, que almacenan granos de almidón, como los encontrados en la raíz de la yuca, el tubérculo de la papa, en granos de cereales, etc. Otros leucoplastos pueden almacenar proteínas, se conocen como proteinoplastos. Los cromoplastos son organelos coloreados, especializados en sintetizar y almacenar pigmentos carotenoides (rojo, anaranjado y amarillo), estos son el origen de los colores de muchos frutos, flores y hojas, por ejemplo, la piel del tomate, la raíz de zanahoria, etc. Los cromoplastos se originan a partir de cloroplastos jóvenes o de cloroplastos maduros, por división.
Los cloroplastos son plastidios que contienen los pigmentos verdes clorofila a y b, así como carotenoides de color anaranjado y xantofilas amarillas, son característicos de los seres fotoautótrofos, que poseen la maquinaria enzimática para transformar la energía solar en energía química, a través de la fotosíntesis. Los cloroplastos son característicos de las células del mesófilo foliar, poseen una doble membrana que los asemeja a las mitocondrias. Tienen una membrana externa y otra interna, el espacio delimitado por la membrana interna está ocupado por un material amorfo, parecido a un gel, rico en enzimas, denominado estroma. Contiene las enzimas que realizan la fijación o reducción del CO2, convirtiéndolo en carbohidratos, como el almidón. La membrana interna de los cloroplastos también engloba un tercer sistema de membranas, que consta de sacos planos llamados tilacoides, en los cuales la energía luminosa se utiliza para oxidar el agua y formar ATP (compuesto rico en energía) y NADPH (poder reductor), usados en el estroma para convertir el CO2 en carbohidratos. En ciertas partes de los cloroplastos, los tilacoides se disponen como monedas apiladas, denominados grana, pero en el estroma permanecen aislados.
Los cloroplastos tienen forma elíptica, con un diámetro de 5 a 10 mm y su número puede variar de 20 a 100 por célula vegetal. Durante la ciclosis se mueven libremente en el citoplasma. Ellos responden directamente a la energía solar, para llevar a cabo la fotosíntesis, orientándose perpendicularmente a los rayos de luz ; sin embargo sí la energía lumínica es muy fuerte , se disponen de tal forma que la radiación incida oblicuamente, recibiendo menos luz.
Los cloroplastos se originan a partir de pro cloroplastos, reacción ésta que es disparada por la luz, que provoca la diferenciación del plastidios, apareciendo los pigmentos y la proliferación de membranas, que origina los tilacoides y grana. Así mismo, en el estroma del cloroplasto se encuentran pequeños pedazos circulares de DNA, dispuestos en doble hélice; parecidos al DNA de las mitocondrias y bacterias. El DNA del cloroplasto regula la síntesis del RNA ribosomal, del RNA de transferencia y de la Ribulosa 1,5 difosfato carboxilasa-oxigenasa (RUBISCO), enzima que cataliza la fijación del CO2 en la fotosíntesis. Sin embargo, la mayoría de las proteínas del cloroplasto, son sintetizadas en el citosol y transportadas al cloroplasto.
Las hojas
Las hojas son expansiones laterales de gran importancia funcional que se presentan en el tallo. Al conjunto de tallo y las hojas se les denomina brote, dado que no existe una separación clara entre ellos.
Hay una estrecha relación entre las hojas y el tallo pues la estructura de los tejidos conductores del tallo y las que se encuentran en el pecíolo y nervio principal de las hojas son semejantes.
Lo mismo puede decirse en cuanto a los tejidos parenquimáticos que se presentan en el córtex del tallo y el mesófilo de la hoja. No obstante, la hoja tiene unas funciones muy específicas: fotosíntesis y transpiración, para las cuales se encuentra muy especializada, tanto en su estructura como en su fisiología.
La forma laminar de la hoja, al aumentar la superficie de contacto con el medio, facilita la captación de energía luminosa y también sus intercambios gaseosos. Las hojas, a diferencia del tallo, no suelen presentar tejidos de reserva y están formadas fundamentalmente por tejidos primarios.
En las hojas llamadas sésiles, el limbo o lamina foliar se une directamente al tallo, mientras que en otras lo hace por medio de un pie llamado pecíolo, el cual se inserta en el tallo por la llamada base foliar.
En la mayoría de las monocotiledóneas y en unas pocas dicotiledóneas (Umbelíferas y Poligonáceas), la base de la hoja presenta un marcado ensanchamiento o vaina que rodea al tallo.
Las hojas son órganos vegetales con crecimiento limitado, normalmente con forma laminar, y estructura dorsiventral (tienen dos caras). Existe una gran diversidad de formas, colores, etc.
Estructura anatómica
La hoja posee una epidermis recubriendo toda la superficie externa. En su interior tiene un tejido parenquimático llamado mesófilo.
La epidermis está provista de estomas, y cubierta de una cutícula cérea cuyo grosor depende de las condiciones ambientales.
En el mesófilo de una hoja bifacial existen dos tipos de parénquima asimilador, organizado de la siguiente manera:
Las hojas Equifaciales son aquellas con exactamente la misma disposición en el haz y el envés, es decir, tiene los tejidos concéntricos unos a otros.
Existen estructuras anatómicas modificadas por las condiciones ambientales. Por ejemplo, en las plantas xerófitas, las hojas se encuentran reducidas debido a que las células están más concentradas para evitar la transpiración. La capa de ceras es más gruesa y una serie de pelos permiten mantener la humedad. Las hojas se pliegan sobre sí mismas, tapando las estomas.
En las hidrófitas, la epidermis prácticamente no tiene ceras. Los mesófilos tienen amplios espacios intercelulares donde se acumula el aire necesario para la respiración.
En cada hoja penetran uno, dos o tres haces vasculares diferentes, los cuales se dividen y constituyen la nervadura o nerviación de la hoja, y los haces del sistema conductor, xilema y floema. En los haces con mayor calibre hay fibras y colénquima, dando mayor consistencia a los órganos foliales. Los nervios principales están formados por un parénquima de transferencia amplio formado por varias capas de células. A esto lo suele recubrir una especie de vainas de células parenquimáticas que puede estar abierta o cerrada.
Morfología Foliar
En el exterior del tallo existen unas protuberancias sin diferenciación llamadas primordios foliares, teniendo crecimiento limitado. Normalmente el crecimiento de hojas se lleva a cabo por meristemas situados en posición apical y otros en posición basal. Y entre ellos pueden aparecer meristemos intercalares y laterales, aumentando la anchura de la hoja.
Habitualmente el meristemo apical cesa su crecimiento junto con el meristemo basal. Pero hay excepciones. existen dos tipos básicos de crecimiento de las hojas.
Además, los meristemos laterales ensanchan la hoja, en las dicotiledóneas esos meristemos laterales son subepidérmico, y pueden ocupar diferentes posiciones. en gramíneas y helechos los meristemos laterales son superficiales.
La hoja se divide en tres partes:
El limbo de las hojas habitualmente tiene simetría dorsiventral, y tiene dos caras, haz y envés o cara ventral y dorsal. El haz suele tener una coloración más oscura que el envés, y esto es resultado de la disposición de los tejidos. En cuánto a su Morfología, hay dos posibilidades:
Morfología del borde foliar: puede tener morfologías muy variadas. En muchos casos los entrantes y salientes se limitan a la periferia de la hoja, pero en otros casos, la hoja es lobulada. Si la incisión sobrepasa la mitad del limbo, la hoja es partida o secta. Dentro de este tipo hay varias divisiones, como las hojas pinnadas (la incisión se dirige hacia el nervio central), o palmadas
Las hojas de tipo compuesto son hojas partidas en las que cada corte llega al nervio central. Cada parte de la hoja se llama foliolo.
Si todos los foliolos parten del extremo del peciolo, sería una hoja palmado-compuesta. O puede ocurrir que los foliolos se inserten a los lados del nervio medio, y en este caso se llama pinnado-compuesta. En este caso el nervio central se llama raquis.
Nerviación de las hojas: se aprecian como unas costillas, más salientes por el envés de la hoja. En la hoja tiene, aparte de la función circulatoria, una función de soporte. Existen tres tipos básicos de nerviaciones:
La más primitiva es la dicótoma bifurcada o flabeliforme. Al final, los nervios llegan a contactar con el borde de la hoja. Por ello, también se le llama «abierta». Se encuentra en bastantes helechos y en un árbol llamado Ginkgo Biloba.
En los dos casos anteriores aparecen uno o varios nervios principales formando una red muy densa, y los nervios de menor tamaño contactan entre sí, y por ello se llaman «cerrados».
La nervadura reticulada (dicotiledóneas y helechos) tiene una o varias venas principales que se ramifican a su vez, pero cuando hay más de una vena principal siempre contactan en un punto. Existen varias modalidades, siendo la craspedódroma la más corriente. La nervadura actinódroma es parecida, pero en hojas palmadas. En la braquidódroma las nervaduras no llegan al borde y entran en contacto entre sí.
En la acródroma existen varias venas principales, las cuales discurren a lo largo de la hoja y se fusionan al llegar al final de la hoja. La campilódroma es parecida, pero más ensanchada y con la convergencia en el extremo.
En la nerviación paralela los nervios principales discurren paralelos unos a otros y siempre están separados en la base.
Existen dos modalidades: el tipo longitudinal estriado y arqueado estriado, y el tipo pinnado estriado.
En cuanto al peciolo, además de ser el punto por donde entran los haces conductores al limbo, actúa como soporte y mantiene al limbo estable. En unas ocasiones, es el tallo el que modifica su posición para exponer la hoja al sol, pero en otras ocasiones, el peciolo tiene un papel fundamental en el movimiento de la hoja. A veces, el peciolo tiene una almohadilla que se llama pulvímulo que actúa de articulación al hincharse o deshincharse. Incluso es posible que en la base de cada foliolo existan pulvímulos.
A veces el movimiento no se debe a la luz, sino que puede ser sensible a los roces, etc. Por ejemplo, la mimosa púdica, en cuanto se la toca, repliega totalmente los foliolos.
Normalmente, la base del peciolo tiene forma arqueada y con la parte abierta hacia el tallo.
Las hojas de una planta no son todas exactamente iguales, sino que varían su morfología según la edad y otros factores. En ocasiones, las hojas de la base son diferentes a las del ápice. A este fenómeno se le denomina anisofilia. Existe una modificación gradual. La heterofilia se da cuando existen en una misma planta hojas muy diferentes, y no hay cambio gradual.
Prefoliación: es la disposición de las hojas en el botón foliar. Previamente forma una especie de joroba que se va ensanchando, y finalmente adopta la forma definitiva de la hoja. La posición de la hoja puede variar de unas plantas a otras. Se representa con un punto la posición del eje del tallo.
Filotaxia o filotaxis es la disposición de las hojas a lo largo de un tallo adulto. En principio se entiende que existen tres tipos básicos:
En cuanto a la disposición verticilada, se cumplen dos premisas:
Los primordios foliares en la yema apical se encuentran en una disposición hexagonal, aunque en planos diferentes. Existen dos parámetros que determinan la posición de los primordios, el tamaño relativo de los primordios en relación al diámetro del cono apical del tallo y La posición relativa de los primordios foliares. Si forman un plano perpendicular al eje principal de la planta, se forman ortósticos.
Existen dos teorías que tratan de explicar la disposición final de las hojas dispersas:
Teoría clásica: es una teoría de naturaleza geométrica. Las hojas con este tipo de disposición se disponen a lo largo del tallo de tal forma que podemos trazar una hélice que une todos los puntos de inserción de las hojas, y en esa hélice vamos a poder situar todas las hojas. A esa hélice se le llama espiga fundamental o generatriz fundamental. La teoría establece que todas las hojas forman entre sí el mismo ángulo de divergencia.
Según esta teoría las hojas forman una serie matemática llamada serie de Fibonacci. Su primer término sería 1/2, el segundo 1/3, el tercero 2/5, 3/8, 5/13,… El numerador indica el número de vueltas que se da al tallo, y el denominador, el número de hojas que encontramos.
Se comprobó que los ángulos de divergencia entre los brotes se encontraban comprendidos entre 137º15´ y 138º. Es decir, el ángulo cambiaba en la zona joven o apical de la planta.
La serie de Fibonacci, como ángulo máximo tiene 137º30´29″. Por tanto, este modelo es bastante válido porque se aproxima a la realidad.
La teoría, sin embargo, tiene una serie de problemas:
La escuela francesa de botánica desarrolló la teoría de las hélices foliares múltiples.
No trata tanto de explicar de forma geométrica la situación, sino que explica el desarrollo de las hojas al crecer el tallo, y tiene muy en cuenta la anatomía de los ápices vegetativos.
Las hojas se insertan a lo largo del tallo distribuidas en series regulares que conforman un número variable de hélices. Va a haber tantas hélices como centros generadores de hojas existan en el ápice del tallo. También sustituye el concepto de ángulo de divergencia por el de contigüidad. Establecen tres tipos diferentes de contigüidad:
Esto se encuentra en consonancia con la disposición de los primordios foliares en el ápice vegetativo del tallo.
Polimorfismo foliar y adaptaciones especiales
La forma de las hojas no sólo depende de la especie, sino que presenta variaciones a lo largo de la misma planta. Las hojas embrionarias son los cotiledones, hojas ya presentes en la semilla, carnosas y repletas de sustancias de reserva. A continuación, se encuentran los catafilos, que aparecen en todas las plantas, de aspecto escamoso y sin clorofila, y se encuentran entre los cotiledones y las primeras hojas de aspecto normal. También se les llama catafilos a las escamas que recubren las yemas enervantes, también llamadas a veces pérulas o tegumentos.
También se encuentran hojas tectrices, en cuya axila aparece una yema lateral. Otro tipo son los profilos, las primeras hojas que se forman cuando empieza a crecer una yema lateral. También hay hipsofilos, hojas de tamaño más reducido que aparecen cerca de las inflorescencias, de aspecto más sencillo que el de las hojas normales de la planta.
Las brácteas son hojas modificadas que se encuentran muy cerca de las flores. La modificación es morfológica, aunque a veces también varía el color. Las flores pueden considerarse hojas modificadas. Se llamarían tépalos a las piezas de la envuelta floral cuando no hay diferencia entre cáliz y corola. A su conjunto se le llama perigonio. Los pétalos son cada una de las piezas corola, y los sépalos cada una de las piezas del cáliz.
Los estambres son hojas modificadas, cuyo conjunto constituye hojas modificadas. El gineceo está formado por carpelos, y también son hojas muy modificadas.
En cuanto a las adaptaciones especiales, se conocen casos de hojas espinosas, como en el acebo y en la encina. Cuando se trata de hojas compuestas, es más complicado, transformándose en espinas la estípula, el raquis, etc.
Hay muchas hojas que se transforman en zarcillos, total o parcialmente. Se encuentran en muchas leguminosas.
Otra modificación son los filodios, hojas con nerviación paralela, y que en realidad corresponden a un peciolo aplanado, como muchas especies de acacia.
También es frecuente encontrar hojas transformadas en órganos de reserva, como cebollas y otras plantas que tienen bulbos basales. También se consideran órganos de reserva a las hojas carnosas de las Crasuláceas.
Las hojas xeromórfica son de pequeño tamaño. Además, tienen una consistencia coriácea. Siempre tienen una gruesa cutícula, y suelen tener pelos entre las estomas hundidos. También repliegan su limbo para encerrar las estomas en su interior. Es muy frecuente que sean Equifaciales.
Las hojas de las plantas insectívoras son de plantas que viven en ambientes muy pobres que se adaptan a capturar pequeños animales, por lo que es indispensable la modificación para captarlos. Ejemplos:
Tejidos parenquimáticos. Clorofílico en empalizada
El parénquima de la hoja vegetativa, el mesófilo, está limitado en sus dos caras por una epidermis. La diferenciación anatómica de dicho parénquima está en correspondencia, con las funciones principales de las hojas, la fotosíntesis de los hidratos de carbono y el desprendimiento de vapor de agua. El parénquima asimilador, rico en cloroplastos, dispuesto bajo la epidermis de la cara superior, se conoce en la mayoría de las hojas de una o varias capas de células cilíndricas dispuestas perpendicularmente a la superficie y recibe a causa de su aspecto en la sección transversal de la hoja, el nombre de parénquima en empalizada; en éste, las células que lo forman, están muy unidas entre sí y son ricas en cloroplastos.
En las hojas con estructura isolateral, el parénquima clorofílico en empalizada se encuentra presente en ambas caras de las hojas.
parénquima cortical, formado por varias células, de forma, tamaño y disposición irregular, guarda dentro de sí varias cámaras intercelulares; también células secretoras llamadas epitélicas, a pesar de ser raras, se presentan en forma abundante en algunas plantas; y la vaina amilífera que es la que limita la corteza del cilindro central o estela.
Periciclo: Meristemo remanente propio de la raíz que se sitúa en la periferia del cilindro vascular. Colabora en la formación del córtex secundario, incluida la peridermis, del cambium vascular, y de las raíces laterales.
Meristemas Secundarios. Tipos. Se hacen cargo de facilitar el crecimiento en grosor.
El cambium vascular: Es un meristemo lateral formado por una monocapa cilíndrica de células situado en aquellos tallos y raíces que van a sufrir engrosamiento secundario. Sus células son de dos tipos: iniciales fusiformes e iniciales radiales, y se dividen en un plano tangencial al tallo o raíz. Las fusiformes dan lugar hacia el interior a células que se diferencian como xilema II y hacia el exterior a células que se diferencian como floema II. Las radiales dan lugar a parénquima. El cambium vascular del tallo se origina de dos formas: Bien a partir de células de procámbium presentes entre el xilema I y el floema I, dando lugar al cambium fascicular, bien por desdiferenciación de células parenquimáticas de los radios medulares, dando lugar al cambium interfascicular.
El cambium fascicular: es el cambium vascular que se forma a partir del procámbium situado entre la xilema y el floema primarios de los haces vasculares.
El cambium interfascicular: Es el cambium vascular que se forma por desdiferenciación de células del parénquima radiomedular, es decir, del parénquima situado entre los haces vasculares .
El cámbium suberógeno o felógeno: Meristemo lateral presente en los tallos y raíces de las plantas con engrosamiento secundario. Es una monocapa cilíndrica de células formada por la desdiferenciación de una capa de parénquima cortical subepidérmico. Por división tangencial estas células generan hacia el interior la felodermis y hacia el exterior el súber (corcho).
| La estela o cilindro central, es la parte central del cuerpo primario del tallo, está formada por el periciclo, los tejidos vasculares, los radios medulares y la médula. El periciclo, se encuentra formando casquete encima del floema. Tejidos Vasculares Primarios. En el tallo primario, la xilema y el floema se agrupan para formar los haces vasculares, que al microscopio se observan como estructuras ovaladas y se les denomina haces colaterales abiertos. |
Los radios medulares: están formados por tejidos parenquimáticos, que se disponen entre los haces vasculares y que comunican la corteza con la médula.
La médula, no es muy común encontrarla en tallos de plantas monocotiledóneas, es la región central y está constituida por tejido parenquimático de almacenamiento o de relleno.
Tejidos vasculares o conductores. Xilema
Es el encargado de transportar de sustancias no elaboradas (savia bruta) a través de todo el cuerpo de las plantas traqueophytas. En las plantas que no poseen tejidos vasculares como las BRIOPHYTAS (musgos), Talophytas (algas, hongos y bacterias) no sucede este proceso.
El xilema es el principal tejido conductor de agua y de sustancias inorgánicas (minerales) y el floema es el tejido conductor de las sustancias alimenticias de una parte de la planta a otra. O sea, los principales tejidos componentes del sistema vascular están representados por la xilema y el floema.
La importancia fisiológica y filogenético del sistema vascular y su destacado papel entre los elementos estructurales del cuerpo de la planta determinó la segregación taxonómica de las plantas provistas de dicho sistema, formando el grupo de las llamadas plantas vasculares.
Xilema:
Al estudiar los meristemos primarios observamos, que el procámbium se localiza dentro del embrión como una banda diferenciada que rompe la continuidad del meristema fundamental o básico, el origina corteza y médula.
El xilema o leño que se diferencia en el cuerpo primario de la planta, se denomina xilema primaria.
Si la planta es de tal naturaleza que después de terminar el crecimiento primario forma tejidos secundarios mediante la actividad del cambium vascular, la xilema formado por este meristemo constituye la xilema secundaria.
El tejido más interno de la estela o cilindro central, adyacente a la médula si la misma está presente, es la xilema primaria.
Este es un tejido compuesto por los siguientes por los siguientes elementos:
Traqueidas
Son las células vasculares donde cada una de ellas queda limitada por membranas transversales primitivas conservando, por lo tanto, la integridad inicial de sus membranas por lo que se les llama vasos cerrados.
Estos vasos son simples células muertas, alargadas y generalmente aguzadas en los extremos, las paredes son delgadas en ciertas regiones alcanzando considerable espesor en otras, producido por la presencia de engrosamientos locales internos.
En la mayoría de las Gimnospermas y algunas Angiospermas menos evolucionadas las traqueidas son los únicos elementos conductores de agua y además sirven como elementos de sostén.
Tráqueas
Son células vasculares donde el enlace de una fila de células superpuestas se produce por resorción de las membranas divisorias primitivas (por lo que también se llaman vasos abiertos).
Por lo antes expuesto, entendemos que una tráquea no es una sola célula, sino un tubo formado por una serie de células cilíndricas muertas, las mayorías de las cuales han disuelto las paredes de los extremos. Estos tubos en la xilema primaria, como las traqueidas, presentan engrosamientos locales internos que pueden ser que pueden ser de los siguientes tipos: anular, espiralado, escalariforme y punteado.
Las tráqueas tienen la pared mucho más engrosada y generalmente lignificada.
Haciendo una comparación entre las traqueidas y las tráqueas vemos que difieren en que las traqueidas poseen células imperforadas, únicamente provistas de pares de puntuaciones en sus membranas, mientras que las membranas de las células que forman las tráqueas están perforadas en ciertas áreas de contacto con otras células. De este modo los elementos de los vasos se unen unos con otros, formando largos tubos continuos, por ello, las traqueidas son células aisladas tubulosas y las tráqueas son tubos relativamente anchos, amplios más raramente estrechos.
Fibras de la xilema primaria (células esclerenquimáticas).
Las fibras de la xilema primaria se derivan del procámbium. Son células alargadas con paredes engrosadas totalmente y por lo general, puntiagudas en los extremos; en algunos casos tienen hasta 1m de largo, pero no es lo común. En ocasiones el engrosamiento de la pared celular es tan grande que la cavidad (lumen) de la célula casi desaparece.
Cuando las fibras han alcanzado su longitud completa y el máximo del engrosamiento de la pared, los protoplastos mueren. Antes de que esto ocurra las paredes se impregnan de lignina.
Esta modificación química de las paredes produce un aumento de la fuerza y dureza del vegetal.
Frecuentemente, las fibras son muy elásticas y se pueden estirar mucho sin perder su capacidad de recuperar su tamaño original.
Células parénquimaticas xilemáticas:
Las células del parénquima están mezcladas con los otros elementos componentes de la xilema primaria. Estas células forman radios medulares por los filos verticales de células vivas en el xilema.
El parénquima asociado con los tejidos vasculares primarios se origina en el procambium.
Las células parénquimaticas xilemáticas pueden ser largas, con contenido variado de sustancias como almidón, grasas, taninos, cristales y otros.
En sus paredes únicamente se encuentran pares de puntuaciones simples.
Xilema secundario
La xilema secundaria también está constituida por varios elementos:
Traqueidas, tráqueas, células parénquimaticas, radios medulares y en ocasiones células secretoras.
Por lo general aparece en las Gimnospermas y en las plantas Dicotiledóneas con crecimiento secundario, y se origina por divisiones tangenciales de las células del cambium vascular, las que se encuentran ordenadas paralelamente a los radios del cuerpo secundario de la planta.
El leño, como también se le denomina, presenta generalmente sus células lignificadas, o sea, con membranas duras y rígidas; en las Dicotiledóneas se componen comúnmente de tres clases distintas de tejidos, con membranas más o menos lignificadas:
Los vasos están representados por traqueidas y tráqueas, las que sirven para el transporte de agua y le proporcionan gran resistencia al tallo. Las traqueidas están formadas por elementos muy amplios y cortos. Estos vasos son del tipo escalariforme o punteado.
Se considera que en las Dicotiledóneas las Traqueidas y las fibras llegan a ser considerablemente más largas que las células de las cuales derivan.
Las fibras leñosas son, por lo general, muy estrechas y largas, con los extremos aguzados y con las membranas gruesas provistas de escasas punteaduras. En dichas fibras el contenido protoplasmático ha sido reabsorbido.
Tílides
En las partes más profundas del leño de muchas plantas, por ejemplo, de Ficus, Vitis, Quercus, se pueden observar pequeñas hernias en el interior de los vasos leñosos que los obstruyen en mayor o en menor grado, los cuales se le denomina tílides. Son formados por abolladuras de las membranas de células parenquimáticas que se hallan siempre circundando los vasos en forma de anillo y que pueden vaciarse completamente o transformarse en elementos reservantes, acumuladores de almidón. Se desarrollan a través de los pares de puntuaciones que relacionan las células parenquimáticas con los elementos traqueales.
Tejidos vasculares o conductores. Floema
Mientras que la xilema es el principal tejido conductor de agua y de sustancias inorgánicas (minerales), el floema es el tejido conductor de las sustancias alimenticias de una parte de la planta a otra.
El segundo tejido de la estela o cilindro central, originado por el procámbium hacia el exterior de la xilema primaria, es el floema primario.
Floema primario. Consta de varios elementos componentes:
Tubos o vasos cribosos:
Sirven para el transporte de las materias orgánicas elaboradas por las plantas, a partir del proceso de fotosíntesis, se componen de células vivientes, dispuestas en series longitudinales y unidos entre sí por medio de poros abiertos que resultan de la perforación local de la membrana divisoria entre dos células procambiales contiguas. Las células de los tubos cribosos presentan protoplastos vivos, muy pobres en plasma. Es un hecho notable que sus núcleos desaparecen muy pronto; presentan leucoplastos, en los que a menudo se observa almidón y, como jugo celular poseen un líquido acuoso que durante el período de asimilación es especialmente rico en sacarosa y a veces también en prótidos.
En general los tubos cribosos son activos únicamente durante un solo período de vegetación. Hacia el final de dicho período, las cribas quedan obstruidas por gruesas acumulaciones de calos, sustancias hialinas que ya mucho antes revestían de una débil película las paredes de los poros. Debido a ello queda interrumpido el intercambio de materia entre los elementos de los tubos cribosos.
Células anexas o acompañantes:
Adyacentes a los tubos cribosos, existen una o más hileras paralelas de células asociadas, que pueden ser de la misma longitud o más cortas que las células que componen los tubos cribosos.
La célula inicial de las anexas tiene el lumen mucho más estrecho que los tubos cribosos, son muy ricas en plasmas, poseen grandes núcleos, a través de un gran número de finas punteaduras y plasmodesmos se comunican con los tubos cribosos y asumen el control de las actividades que ocurren en el plasma de éstos.
Células parénquimaticas:
El floema contiene, además de las células acompañantes, células parenquimáticas, en cantidad variable, y son las encargadas del almacenamiento de grasa, almidón y otros materiales orgánicos alimenticios y acumulaciones de taninos y resinas.
Estas células son alargadas y están orientadas de forma que sus ejes longitudinales son paralelos a la dirección longitudinal del tejido vascular.
Fibras:
Pueden alcanzar gran longitud. Sus células forman membranas secundarias después de complementar su alargamiento. En algunas plantas las fibras están lignificadas típicamente, en otras no. Las puntuaciones son por lo general simples; también pueden presentarse rebordeadas o areoladas.
Tejidos vasculares de origen secundario:
En la mayoría de las Dicotiledóneas y Gimnospermas, los tejidos vasculares primarios en el tallo están dispuestos en forma de haces agrupados circularmente; con la xilema hacia el interior y el floema hacia el exterior. Como ya se ha dicho, una parte de las células del procámbium permanecen meristemáticas aún después de completarse el crecimiento primario, desarrollándose por consiguiente el cambium vascular en forma de un cilindro ininterrumpido entre los haces vasculares vecinos.
Este cilindro discontinuo pronto se hace continuo ya que las capas de parénquima entre los haces cercanos manifiestan actividad meristemática, formando el cambium interfascicular. De esta manera, el cambium vascular en el tallo está constituido de porciones alternas de cambium fascicular e interfascicular.
El cambium fascicular por repetidas divisiones mitóticas de sus células forma leño hacia la parte interna y liber hacia la parte externa, mientras que el cambium interfascicular forma además parénquima radiomedular. En algunas Dicotiledóneas herbáceas con crecimiento secundario, el cambium interfascicular produce solamente fibras o sólo parénquima esclerotizado en el lado de la xilema (Medicago), o bien el crecimiento secundario puede ser tan pequeño que queda limitado a los haces vasculares (Cucúrbita).
En la raíz de Dicotiledóneas y Gimnospermas, la parte principal del cambium vascular, se forma entre la xilema primaria y el floema en la estela. Cuando se produce crecimiento secundario en ella, una capa de células de procámbium se forma entre la xilema y el floema, se conserva permanente embriónico y no diferenciado, representando una parte del cambium vascular de la raíz. El resto forma poco después las células parénquimaticas del periciclo. Este cambium vascular se presenta primero el borde interno de los cordones floemáticas.
Floema secundario
El líber secundario se forma por la actividad del cambium hacia el interior de los cordones liberianos primarios y constituyen la parte principal de la corteza secundaria; está formado por diversos elementos que se superponen de manera regular. Estos elementos son:
Los tubos cribosos, con placas perforadas, y en las Dicotiledóneas acompañadas de células anexas ricas en plasma.
El parénquima liberiano, constituido de células de membranas sutiles, que se llenan de almidón o de sustancias proteicas, (como en las Gimnospermas).
Las fibras esclerenquimáticas (por lo general muertas).
Se puede observar, además, esclereidas, radios medulares, células crístaliferas, secretoras, tubos laticíferos y células mucilaginíferas.
En ciertas Dicotiledóneas se presenta una gran diversidad de tipos de ordenación en sus células, presentándose las fibras dispuestas en bandas tangenciales, alternando más o menos regularmente con bandas de tubos cribosos, que por lo general forman conductos longitudinales ininterrumpidos y ramificados que van desde las raíces a las hojas.
Estos tubos limitan al que los cordones de parénquima liberiano, con los radios medulares, también longitudinales, los que constituyen la prolongación radical de los radios medulares del leño.
Tejidos vasculares o conductores. Haces
Una porción del sistema vascular primario del tallo o de la hoja, constituye un cordón o haz vascular.
El floema y la xilema se hallan asociados no sólo en su conjunto, sino también por lo regular dentro de sus partes, es decir, en los haces vasculares. Las diferentes maneras de disponerse los tejidos vasculares dentro de los haces ha permitido clasificarlos en distintos tipos:
Haces colaterales abiertos:
Son aquellos que conservan algunas células meristemáticas (o procambiales) entre el floema (líber) y el xilema (leño), productoras de elementos secundarios con origen primario.
Estas células procambiales constituyen una zona generatriz, integrada por células meristemáticas situadas entre el floema y la xilema, la cual por repetidas divisiones tangenciales de sus células produce xilema secundaria hacia la parte interna y floema secundario hacia el exterior. Poseen cambium en general las Gimnospermas y las Dicotiledóneas y su actividad se debe al crecimiento secundario de los mismos. El cambium que se halla entre la xilema y el floema se llama cambium fascicular.
Haces colaterales cerrados:
Se consideran cerrados los haces donde el floema y la xilema están en contacto inmediato, incapaces de modificar su estructura primitiva. Este tipo de haz es característico de las plantas monocotiledóneas. En estos tipos de haces todas las células del procámbium se diferencian en elementos de la xilema y el floema y ninguno permanece en estado meristemático como aquellos del cambium fascicular de las Gimnospermas y Dicotiledóneas. Debido a la ausencia del cambium, la mayoría de las monocotiledóneas no experimentan crecimiento en grosor.
En los haces conductores existen generalmente vainas de tejidos mecánicos en el lado exterior del floema y, a veces también en lado interno del haz, por fuera de la xilema; estas vainas en sección transversal aparecen como extractos semilunares de esclerénquima. Son pues, incompletos en los haces abiertos. A cada lado del haz, es decir, junto al límite entre la xilema y el floema, la vaina se convierte en una banda de células parenquimáticas más débilmente engrosadas, que permiten el intercambio de agua y materias alimenticias entre el haz y el parénquima circundante.
Haces bicolaterales:
Se presentan en determinadas familias de plantas como Cucurbitáceas y Solanáceas; estas poseen dos zonas de floema: una en posición normal, como en el haz colateral y la otra más hacia adentro que el leño y menos desarrollado que la primera.
Haces concéntricos:
En algunas Pteridofitas y Monocotiledóneas el tipo de haz es denominado concéntrico; los hay de dos clases atendiendo a la posición de los tejidos vasculares. Con la xilema en la parte externa rodeando el floema, (tipo frecuente en los rizomas de las Monocotiledóneas), llamándose por ello concéntrico perixilemático. Y con la xilema en la parte central y el floema al exterior (frecuentemente se presenta en los helechos, por ejemplo, en el género Pteris), se designa con el nombre de concéntrico perifloemático.
Haces radiales:
Se encuentra en la raíz y es característico de este órgano. Este haz conductor radial comprende varios cordones xilemáticas y floemáticas, que se disponen como los radios de una rueda, alternando los de una clase con lo de la otra. El conjunto de los cordones xilemáticos de ordinario aplanado lateralmente, presenta pues forma estrellada en la sección transversal. Los entrantes comprendidos entre las láminas leñosas están ocupados por los cordones cribosos, que quedan separados de los cordones xilemáticos por una o varias capas de células parenquimáticas.
En los haces radiales, la formación de todos los elementos de los cordones xilemáticos y floemáticos avanzan progresivamente de la periferia del haz hacia el centro del mismo, en la porción más externa quedan por ello las células primiciales o primarias (el protoxilema y protofloema); les sigue hacia el interior los elementos diferenciados más tarde (metaxilema y metafloema). Estos haces son típicos de raíces; en el vástago se presenta raramente, por ejemplo, en algunos helechos. El número de radios xilemáticos varía de unas especies a otras.
El microscópio
Microscopio, cualquiera de los distintos tipos de instrumentos que se utilizan para obtener una imagen aumentada de objetos minúsculos o detalles muy pequeños de los mismos.
Microscopio óptico
El tipo de microscopio más utilizado es el microscopio óptico, que se sirve de la luz visible para crear una imagen aumentada del objeto. El microscopio óptico más simple es la lente convexa doble con una distancia focal corta. Estas lentes pueden aumentar un objeto hasta 15 veces. Por lo general se utilizan microscopios compuestos, que disponen de varias lentes con las que se consiguen aumentos mayores. Algunos microscopios ópticos pueden aumentar un objeto por encima de las 2.000 veces.
El microscopio compuesto consiste en dos sistemas de lentes, el objetivo y el ocular, montados en extremos opuestos de un tubo cerrado. El objetivo está compuesto de varias lentes que crean una imagen real aumentada del objeto examinado. Las lentes de los microscopios están dispuestas de forma que el objetivo se encuentre en el punto focal del ocular. Cuando se mira a través del ocular se ve una imagen virtual aumentada de la imagen real. El aumento total del microscopio depende de las longitudes focales de los dos sistemas de lentes.
El equipamiento adicional de un microscopio consta de un armazón con un soporte que sostiene el material examinado y de un mecanismo que permite acercar y alejar el tubo para enfocar la muestra. Los especímenes o muestras que se examinan con un microscopio son transparentes y se observan con una luz que los atraviesa, y se suelen colocar sobre un rectángulo fino de vidrio. El soporte tiene un orificio por el que pasa la luz. Bajo el soporte se encuentra un espejo que refleja la luz para que atraviese el espécimen. El microscopio puede contar con una fuente de luz eléctrica que dirige la luz a través de la muestra.
La fotomicrografía, que consiste en fotografiar objetos a través de un microscopio, utiliza una cámara montada por encima del ocular del microscopio. La cámara suele carecer de objetivo, ya que el microscopio actúa como tal. El término microfotografía, utilizado a veces en lugar de fotomicrografía, se refiere a una técnica de duplicación y reducción de fotografías y documentos a un tamaño minúsculo para guardarlos en un archivo.
Los microscopios que se utilizan en entornos científicos cuentan con varias mejoras que permiten un estudio integral del espécimen. Dado que la imagen de la muestra está ampliada muchas veces e invertida, es difícil moverla de forma manual. Por ello los soportes de los microscopios científicos de alta potencia están montados en una plataforma que puede moverse con tornillos micrométricos. Algunos microscopios cuentan con soportes giratorios. Todos los microscopios de investigación cuentan con tres o más objetivos montados en un cabezal móvil que permite variar la potencia de aumento.
Microscopios ópticos especiales
Hay diversos microscopios ópticos para funciones especiales. Uno de ellos es el microscopio estereoscópico, que no es sino un par de microscopios de baja potencia colocados de forma que convergen en el espécimen. Estos instrumentos producen una imagen tridimensional.
El microscopio de luz ultravioleta utiliza el rango ultravioleta del espectro luminoso en lugar del rango visible, bien para aumentar la resolución con una longitud de onda menor o para mejorar el detalle absorbiendo selectivamente distintas longitudes de onda de la banda ultravioleta. Dado que el vidrio no transmite las longitudes de onda más cortas de la luz ultravioleta, los elementos ópticos de estos microscopios están hechos con cuarzo, fluorita o sistemas de espejos aluminizados. Además, dado que la radiación ultravioleta es invisible, la imagen se muestra con fosforescencia (véase Luminiscencia), en fotografía o con un escáner electrónico. El microscopio de luz ultravioleta se utiliza en la investigación científica.
El microscopio petrográfico o de polarización se utiliza para identificar y estimar cuantitativamente los componentes minerales de las rocas ígneas y las rocas metamórficas. Cuenta con un prisma de Nicol u otro tipo de dispositivo para polarizar la luz que pasa a través del espécimen examinado (véase Óptica: Polarización de la luz). Otro prisma Nicol o analizador determina la polarización de la luz que ha pasado a través del espécimen. El microscopio tiene un soporte giratorio que indica el cambio de polarización acusado por el espécimen.
El microscopio en campo oscuro utiliza una luz muy intensa en forma de un cono hueco concentrado sobre el espécimen. El campo de visión del objetivo se encuentra en la zona hueca del cono de luz y sólo recoge la luz que se refleja en el objeto. Por ello las porciones claras del espécimen aparecen como un fondo oscuro y los objetos minúsculos que se están analizando aparecen como una luz brillante sobre el fondo. Esta forma de iluminación se utiliza para analizar elementos biológicos transparentes y sin manchas, invisibles con iluminación normal.
El microscopio de fase ilumina el espécimen con un cono hueco de luz, como en el microscopio en campo oscuro. Sin embargo, en el microscopio de fase el cono de luz es más estrecho y entra en el campo de visión del objetivo, que contiene un dispositivo en forma de anillo que reduce la intensidad de la luz y provoca un cambio de fase de un cuarto de la longitud de onda. Este tipo de iluminación provoca variaciones minúsculas en el índice de refracción de un espécimen transparente, haciéndolo visible. Este tipo de microscopio es muy útil a la hora de examinar tejidos vivos, por lo que se utiliza con frecuencia en biología y medicina.
Entre los microscopios avanzados se encuentran el microscopio de campo cercano, con el que pueden verse detalles algo menores a la longitud de onda de la luz. Se hace pasar un haz de luz a través de un orificio diminuto y se proyecta a través del espécimen a una distancia equivalente a la mitad del diámetro del orificio, formando una imagen completa.
Microscópio electrónico
La potencia amplificadora de un microscopio óptico está limitada por la longitud de onda de la luz visible. El microscopio electrónico utiliza electrones para iluminar un objeto. Dado que los electrones tienen una longitud de onda mucho menor que la de la luz pueden mostrar estructuras mucho más pequeñas. La longitud de onda más corta de la luz visible es de alrededor de 4.000 Ángstrom (1 ángstrom es 0,0000000001 metros). La longitud de onda de los electrones que se utilizan en los microscopios electrónicos es de alrededor de 0,5 ángstroms.
Todos los microscopios electrónicos cuentan con varios elementos básicos. Disponen de un cañón de electrones que emite los electrones que chocan contra el espécimen, creando una imagen aumentada. Se utilizan lentes magnéticas para crear campos que dirigen y enfocan el haz de electrones, ya que las lentes convencionales utilizadas en los microscopios ópticos no funcionan con los electrones. El sistema de vacío es una parte relevante del microscopio electrónico. Los electrones pueden ser desviados por las moléculas del aire, de forma que tiene que hacerse un vacío casi total en el interior de un microscopio de estas características. Por último, todos los microscopios electrónicos cuentan con un sistema que registra o muestra la imagen que producen los electrones.
Hay dos tipos básicos de microscopios electrónicos: el microscopio electrónico de transmisión (Transmission Electron Microscope, TEM) y el microscopio electrónico de barrido (Scanning Electron Microscope, SEM). Un TEM dirige el haz de electrones hacia el objeto que se desea aumentar. Una parte de los electrones rebotan o son absorbidos por el objeto y otros lo atraviesan formando una imagen aumentada del espécimen. Para utilizar un TEM debe cortarse la muestra en capas finas, no mayores de un par de miles de ángstroms. Se coloca una placa fotográfica o una pantalla fluorescente detrás del objeto para registrar la imagen aumentada. Los microscopios electrónicos de transmisión pueden aumentar un objeto hasta un millón de veces.
Un microscopio electrónico de barrido crea una imagen ampliada de la superficie de un objeto. No es necesario cortar el objeto en capas para observarlo con un SEM, sino que puede colocarse en el microscopio con muy pocos preparativos. El SEM explora la superficie de la imagen punto por punto, al contrario que el TEM, que examina una gran parte de la muestra cada vez. Su funcionamiento se basa en recorrer la muestra con un haz muy concentrado de electrones, de forma parecida al barrido de un haz de electrones por la pantalla de una televisión. Los electrones del haz pueden dispersarse de la muestra o provocar la aparición de electrones secundarios. Los electrones perdidos y los secundarios son recogidos y contados por un dispositivo electrónico situado a los lados del espécimen. Cada punto leído de la muestra corresponde a un píxel en un monitor de televisión. Cuanto mayor sea el número de electrones contados por el dispositivo, mayor será el brillo del píxel en la pantalla. A medida que el haz de electrones barre la muestra, se presenta toda la imagen de la misma en el monitor. Los microscopios electrónicos de barrido pueden ampliar los objetos 100.000 veces o más. Este tipo de microscopio es muy útil porque, al contrario que los TEM o los microscopios ópticos, produce imágenes tridimensionales realistas de la superficie del objeto.
Se han desarrollado otros tipos de microscopios electrónicos. Un microscopio electrónico de barrido y transmisión (Scanning Trasnmission Electron Microscope, STEM) combina los elementos de un SEM y un TEM, y puede mostrar los átomos individuales de un objeto. El microanalizador de sonda de electrones, un microscopio electrónico que cuenta con un analizador de espectro de rayos X, puede analizar los rayos X de alta energía que produce el objeto al ser bombardeado con electrones. Dado que la identidad de los diferentes átomos y moléculas de un material se puede conocer utilizando sus emisiones de rayos X, los analizadores de sonda de electrones no sólo proporcionan una imagen ampliada de la muestra, como hace un microscopio electrónico, sino que suministra también información sobre la composición química del material.
Microscópio de sonda de barrido
En los microscopios de sonda de barrido se utiliza una sonda que recorre la superficie de una muestra, proporcionando una imagen tridimensional de la red de átomos o moléculas que la componen. La sonda es una afilada punta de metal que puede tener un grosor de un solo átomo en su extremo. Un tipo importante de microscopio de sonda de barrido es el microscopio de túnel de barrido (siglas en inglés de Scanning Tunelling Microscope, STM) desarrollado en 1981. Este microscopio utiliza un fenómeno de la física cuántica, denominado efecto túnel, para proporcionar imágenes detalladas de sustancias conductoras de electricidad. La sonda se coloca a una distancia de pocos ángstroms de la superficie del material y se aplica un voltaje pequeño entre la superficie y la sonda. A causa de la poca distancia entre el material y la sonda algunos electrones se escapan a través del hueco, generando una corriente. La magnitud de la corriente del efecto túnel depende de la distancia entre la superficie y la sonda. El flujo de corriente es mayor cuando la sonda se acerca al material y disminuye cuando se aleja. A medida que el mecanismo de barrido mueve la sonda por encima de la superficie, se ajusta de modo automático la altura de la sonda para mantener constante la corriente del efecto túnel. Estos ajustes minúsculos permiten dibujar las ondulaciones de la superficie. Después de muchas pasadas hacia adelante y hacia atrás se utiliza una computadora para crear una representación tridimensional del material.
Otro tipo de microscopio de sonda de barrido es el microscopio de fuerza atómica (Atomic Force Microscope, AFM), que no emplea la corriente de efecto túnel y que por lo tanto puede utilizarse también en materiales no conductores. A medida que la sonda se mueve a lo largo de la superficie de la muestra los electrones de la sonda de metal son repelidos por las nubes electrónicas de los átomos de la misma. La altura de la sonda se ajusta de modo automático para mantener constante la fuerza recibida. Un sensor registra el movimiento ascendente y descendente de la sonda y entrega la información a una computadora, que a su vez la utiliza para dibujar una imagen tridimensional de la superficie del espécimen.
Partes más importantes de un microscopio óptico
Ocular (Ocular): Qué el utilizador mira a través para examinar el espécimen. La potencia del ocular multiplicada por la potencia objetiva iguala la ampliación total (es decir 10X objetivo 10X del ocular X = 100X. El espécimen se ha agrandado 100 veces).
Cuerpo: El microscopio puede venir como un monocular, (un ocular y tubo), o binocular, (dos oculares y tubos). Si es un binocular generalmente solamente un tubo del ocular será ajustable mientras que el otro es fijo.
Objetivos: El objetivo es la lente más importante del microscopio para producir una imagen clara de la alta resolución. El objetivo tiene varias funciones importantes. Debe recolectar la luz que viene de cada uno de las varias partes o puntas del espécimen. Debe tener la capacidad de reconstituir la luz que viene de las varias puntas del espécimen en las varias puntas correspondientes de la imagen. El objetivo se debe construir de modo que sea enfocado cerca bastante al espécimen para proyectar una imagen magnificada, verdadera para arriba en el tubo del cuerpo.
Etapa mecánica: Un dispositivo para llevar a cabo diapositivas con seguridad con las perillas con estrías separadas para mover la diapositiva desde frente a la parte posterior (norte y al sur) o de lado a lado (este y al oeste). Estas perillas pueden estar en los ejes separados o en un eje coaxial. Pueden enderezar o zurdo.
Condensador del sub-stage: Se cabe debajo entre de la etapa del microscopio, la lámpara que ilumina y el espécimen. La abertura del condensador y el enfocarse apropiado del condensador son de importancia crítica en realizar la capacidad máxima de la lente objetiva en uso. Asimismo, el uso apropiado del diafragma ajustable del diafragma de la abertura (incorporado en el condensador) es también importante para asegurar la iluminación apropiada y contraste. La apertura y el cierre del diafragma del diafragma controla el ángulo de los rayos de la iluminación que pasan a través del condensador, a través del espécimen y entonces en la lente objetiva.
Controles del foco de Fine/Coarse: También suele llamárseles tornillos macrométrico y micrométrico, en ambas caras del soporte del microscopio hay dos conjuntos de perillas del ajuste. La perilla del ajuste aproximado para movimientos que se enfocan incrementales más grandes y la perilla del ajuste fino para movimientos que se enfocan incrementales más pequeños. Las perillas del ajuste sirven para traer el objetivo y el espécimen más cercano juntas o para engendrar aparte. En la mayoría de los microscopios las perillas del ajuste levantan o bajan la etapa; en algunos microscopios las perillas levantan o bajan el tubo el microscopio o el pedazo de la nariz.
Iluminador (Estándar): Esta parte proporciona a la iluminación requerida para realizar cualquier función con el microscopio. Es básicamente una fuente de alimentación electrónica que proporciona a electricidad a la fuente de la lámpara. Esta fuente de la lámpara podía ser una lámpara estándar del tungsteno o lámpara del halógeno. **time-out** ventaja halógeno tipo lámpara ser más luz para publicar salida vatiaje y en bajo nivel iluminación luz no tender para dar vuelta amarilla a medida que estándar tungsteno lámpara hacer.
Cubierta Del Iluminador (Estándar): Aquí es adonde el elemento electrónico asociado al iluminador se pone. Algunos microscopios más viejos tienen transformadores separados del control y el único interior de la cubierta es la lámpara, ensamblaje del socket, condensador del campo y el diafragma del diafragma del campo.
Diafragma del campo Condenser/Iris: Esta pieza del microscopio contiene la lente de condensador de la lámpara y el diafragma del diafragma del campo. Este diafragma controla el área del círculo de la luz que ilumina el espécimen.
Tejidos vasculares o conductores. Haces
Una porción del sistema vascular primario del tallo o de la hoja, constituye un cordón o haz vascular.
El floema y la xilema se hallan asociados no sólo en su conjunto, sino también por lo regular dentro de sus partes, es decir, en los haces vasculares. Las diferentes maneras de disponerse los tejidos vasculares dentro de los haces ha permitido clasificarlos en distintos tipos:
Haz colateral abierto:
Es aquel que conserva algunas células meristemáticas (o procambiales) entre el floema (líber) y la xilema (leño), productoras de elementos secundarios con origen primario.
Estas células procambiales constituyen una zona generatriz, integrada por células meristemáticas situadas entre el floema y la xilema, la cual por repetidas divisiones tangenciales de sus células produce xilema secundaria hacia la parte interna y floema secundario hacia el exterior. Poseen cambium en general las Gimnospermas y las Dicotiledóneas y su actividad se debe al crecimiento secundario de los mismos. El cambium que se halla entre la xilema y el floema se llama cambium fascicular.
Haz colateral cerrado:
Se considera cerrado el haz donde el floema y la xilema están en contacto inmediato, incapaces de modificar su estructura primitiva. Este tipo de haz es característico de las plantas monocotiledóneas. En estos tipos de haces todas las células del procámbium se diferencian en elementos de la xilema y el floema y ninguno permanece en estado meristemático como aquellos del cambium fascicular de las Gimnospermas y Dicotiledóneas. Debido a la ausencia del cambium, la mayoría de las monocotiledóneas no experimentan crecimiento en grosor.
En los haces conductores existen generalmente vainas de tejidos mecánicos en el lado exterior del floema y, a veces también en lado interno del haz, por fuera de la xilema; estas vainas en sección transversal aparecen como extractos semilunares de esclerénquima. Son pues, incompletos en los haces abiertos. A cada lado del haz, es decir, junto al límite entre la xilema y el floema, la vaina se convierte en una banda de células parenquimáticas más débilmente engrosadas, que permiten el intercambio de agua y materias alimenticias entre el haz y el parénquima circundante.
Haz bicolateral:
Se presentan en determinadas familias de plantas como Cucurbitáceas y Solanáceas; estas poseen dos zonas de floema: una en posición normal, como en el haz colateral y la otra más hacia adentro que el leño y menos desarrollado que la primera.
Haz concéntrico:
En algunas Pteridofitas y Monocotiledóneas el tipo de haz es denominado concéntrico; los hay de dos clases atendiendo a la posición de los tejidos vasculares. Con la xilema en la parte externa rodeando el floema, (tipo frecuente en los rizomas de las Monocotiledóneas), llamándose por ello concéntrico perixilemático. Y con la xilema en la parte central y el floema al exterior (frecuentemente se presenta en los helechos, por ejemplo, en el género Pteris), se designa con el nombre de concéntrico perifloemático.
Haz radial:
Se encuentra en la raíz y es característico de este órgano. Este haz conductor radial comprende varios cordones xilemáticos y floemáticos, que se disponen como los radios de una rueda, alternando los de una clase con lo de la otra. El conjunto de los cordones xilemáticos de ordinario aplanado lateralmente, presenta pues forma de estrellados en la sección transversal. Los entrantes comprendidos entre las láminas leñosas están ocupados por los Cordones cribosos, que quedan separados de los cordones xilemáticos por una o varias capas de células parenquimáticas.
En el haz radial, la formación de todos los elementos de los cordones xilemáticos y floemáticos avanzan progresivamente de la periferia del haz hacia el centro del mismo, en la porción más externa quedan por ello las células primiciales o primarias (el protoxilema y protofloema); les sigue hacia el interior los elementos diferenciados más tarde (metaxilema y metafloema). Estos haces son típicos de raíces; en el vástago se presenta raramente, por ejemplo, en algunos helechos. El número de radios xilemáticos varía de unas especies a otras.
Parénquima reservante.
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas, presenta funciones metabólicas grandes vacuolas.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
De acuerdo con su diferenciación funcional se puede distinguir: parénquima de relleno, clorofílicos, reservantes, acuífero y aerífero.
Tejidos parenquimáticos. Relleno
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
La función principal del parénquima depende del tipo de especialización que presente, y podría mencionarse el almacenamiento y producción de sustancias alimenticias, transporte de sustancias célula a célula, producir raíces adventicias, reparar daños de la corteza, excreción y secreción, etc.
También, en muchos órganos (ej. hojas con estructura isolateral, tallos y frutos), tejidos especializados, como los vasculares, (floema), aparece en forma de tejidos parenquimáticos pocos diferenciados, en los que actúa como un tejido de relleno, brindándole solidez al órgano o tejido en cuestión.
En el cuerpo de la planta el parénquima fundamental constituye la masa en la que se encuentran incluidos todos los demás tejidos. Gracias a la turgencia de sus células sirve para dar solidez general al cuerpo vegetativo, de hecho, etimológicamente, la palabra parénquima significa tejido que está en medio (tejido de relleno).
Este tejido se carácteriza por:
Tallos
Es la porción del eje de la planta que sostiene las hojas, se desarrolla a partir de la plúmula del embrión y se caracteriza por la presencia de nudos y entrenudos. En cada nudo se encuentra una o varias hojas, hojas modificadas o cicatrices de hojas. La región entre dos nudos constituye un entrenudo.
En el ápice y en las axilas (de una hoja o superior a la cicatriz de ésta) se encuentran las yemas. Algunas yemas axilares pueden ser modificadas, las yemas son estructuras no desarrollada de los tallos. Se clasifican por su posición u origen (terminales, axilares, accesorias o adventicias); por su actividad (activas o latentes) y por su contenido (foliar, floral o mixto).
La función del tallo está relacionada con el transporte de agua y nutrientes a través de la planta, además sirve como sitio de reserva de asimilados y sostén de las partes aéreas.
Clases de tallos:
Desde el tiempo de los antiguos botánicos griegos como Teofrasto y hasta el siglo XVII, la división clásica de las plantas establecía, según la estructura del tallo, tres grupos: hierbas, arbustos y árboles.
Los tallos pueden ser epigeos, si se desarrollan en el aire, hipogeos o subterráneos, cuando lo hacen enterrados; en este caso se les da los nombres de rizomas, tubérculos o bulbos.
Anatomía.
Los meristemos apicales son los generadores de los primeros tejidos del tallo, de la raíz y de los apéndices (hojas). Los tejidos primarios son estructural y funcionalmente íntegros, al menos temporalmente, y conforman el cuerpo primario de la planta.
Los tejidos secundarios constituyen, el cuerpo y pueden ser agregados más tarde; son tejidos suplementarios, reemplazan o refuerzan estructural y funcionalmente ciertos tipos de tejidos primarios como la epidermis o para la protección, y los tejidos vasculares primarios, xilema y floema para la conducción.
El crecimiento primario constituye el cuerpo de la mayoría de monocotiledóneas y pteridofitas. El crecimiento secundario se agrega al crecimiento primario en gimnospermas y muchas dicotiledóneas. El crecimiento secundario se origina en el cambium felógeno que incrementan el grosor del eje.
Los tejidos permanentes del tallo son:
Las células parenquimáticas fotosintéticas contienen cloroplastos, algunas tienen leucoplastos y otras sirven de reserva de diferentes materiales como azucares, grasas y almidones.
Colénquima y esclerénquima: El esclerénquima está constituido por células con paredes secundarias, engrosadas, lignificadas o no. La función básica es mecánica. En el esclerénquima existen dos tipos reconocidos comúnmente de células, las fibras y las esclereidas. Las células de esclerénquima a diferencia de las de la colénquima contienen un bajo porcentaje de agua y en la madurez no tienen protoplasma floema y xilema:
1.- Sistema vascular primario: Los tejidos vasculares primarios del tallo corresponden al xilema y floema primarios; estos constituyen el esqueleto básico.
Difieren entre las espermatofitas en que en las gimnospermas y en la mayoría de las dicotiledóneas el cuerpo vascular primario consiste de un cilindro continuo que envuelve la medula, esto es la porción central del tallo. En las dicotiledóneas herbáceas está constituido por un número de haces estrechos y separados que forman un anillo.
La xilema y el floema de los tallos de monocotiledóneas nunca están dispuestos en capas continuas, sino en haces vasculares individuales, esparcidos a través del tallo en forma desorganizada, más que en círculo como ocurre en las dicotiledóneas que tiene sus tejidos vasculares en haces separados. El procámbium es el tejido que origina la xilema y el floema primarios. El procámbium más temprano aparece como bandas aisladas muy estrechas en el ápice del tallo; se continua hacia las zonas más desarrolladas y diferencia el tejido vascular primario que se desarrolla en forma acropetal. Las primeras células de un haz vascular que maduran corresponden al floema, luego le siguen las primeras de la xilema.
2.- Sistema vascular secundario: La disposición del tejido vascular secundario no tiene relación con el existente tejido vascular primario, la cual puede ser en forma de cilindro completo. Sin embargo, la cantidad y disposición de los tejidos secundarios, especialmente la de la xilema, puede variar desde un cilindro completo del mismo grosor, como en árboles, hasta bandas separadas como en los tallos herbáceos de algunas dicotiledóneas, caso de Cucúrbita sp.
La xilema es un tejido constituido de varios tipos de células: células no vivas relacionadas con el transporte de agua y solutos, las fibras que ayudan al fortalecimiento del cuerpo de la planta y células del parénquima para la reserva de agua.
Dentro de los elementos traqueales se diferencian dos tipos de células: traqueidas y tráqueas. Las traqueidas son células no perforadas; las áreas de contacto presentan solamente punteaduras dobles. Las tráqueas o vasos, muestran perforaciones en sus extremos, sin embargo, existen también perforaciones en las paredes laterales.
Al completar los engrosamientos y la lignificación de la pared, el protoplasto desaparece en las células de la xilema. El floema es un tejido compuesto de elementos cribosos, y células parenquimáticas especializadas llamadas anexas. Los elementos cribosos se caracterizan por presentar cribas en sus paredes, protoplasto sin núcleos y pared primaria constituida principalmente por celulosa. La función del floema es el transporte del producto de la fotosíntesis.
Tejidos parenquimáticos. Reservante
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancia de reserva. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
El parénquima reservante, se encuentra en los órganos reservantes como en los cotiledones de las semillas, la médula, en la corteza de los tallos y raíces, los radios medulares, siendo, su función la de almacenar y transportar sustancias de reserva. Sus células tienen membranas sutiles, con pequeños espacios intercelulares. Almacena carbohidratos, proteínas y lípidos.
Las raíces
Órgano de las plantas superiores, casi siempre subterráneo, que desempeña varias funciones, entre ellas absorber y conducir agua y minerales disueltos, acumular nutrientes y sujetar la planta al suelo.
La raíz se diferencia del tallo por su estructura, por el modo en que se forma y por la falta de apéndices, como yemas y hojas. La primera raíz de la planta, llamada radícula, se alarga cuando germina la semilla y forma la raíz primaria.
Las raíces que se ramifican a partir de la primaria se llaman secundarias. En muchas plantas, la raíz primaria se llama pivotante, es mucho mayor que las secundarias y alcanza mayor profundidad en el suelo.
La remolacha la zanahoria son ejemplos característicos de plantas con gruesas raíces pivotantes. Algunas especies con raíces de este tipo son difíciles de trasplantar, porque la rotura de la raíz primaria determina la pérdida de casi todo el sistema radicular y la muerte de la planta.
Las raíces que brotan de los tallos se llaman adventicias. Se ven estas formaciones cerca de la base del tallo ya que se originan de cualquier parte menos de la radícala, un ejemplo de estas raíces es la que produce la yedra venenosa, estas raíces cortas tienen estructuras terminales planas que permiten fijarse.
Cuando brotan de puntos más altos, las raíces adventicias se llaman aéreas, y ayudan a sujetar la planta, como se observa en el banano, el mangle y ciertas orquidáceas.
Las plantas parásitas usan unas modificaciones de la raíz llamadas haustorios un ejemplo de esto es la cuscuta, se fija al tejido vascular de sus hospederos gracias a estas modificaciones que les permite suprimir el abastecimiento de agua y nutrientes.
Al contrario de las parásitas, estas no obtienen beneficio de la planta que lo sostiene. Además, existen plantas hemiparásitas, es decir que toman sus nutrientes del organismo huésped, y producen parte de su propio alimento.
Las raíces, de acuerdo al medio donde se desarrollan, pueden ser
Por su duración las clasificamos en
Composición
La raíz está formada por tres tipos de tejido: epidermis, o capa superficial; tejido fundamental o córtex; y estela o cilindro vascular, situado en el centro.
Algunas células de la epidermis se modifican para desempeñar la función de absorción; de ellas parten largas proyecciones tubulares llamadas pelos radicales que se sujetan a las partículas del suelo. El agua absorbida por los pelos radicales atraviesa el córtex, zona dedicada al almacenamiento de agua y nutrientes, y penetra en el cilindro vascular, que la conduce hacia el tallo.
La organización del cilindro vascular de la raíz es muy distinta de la disposición del tejido vascular del tallo. En éste, xilema y floema se agrupan en haces vasculares; por el contrario, la raíz tiene un núcleo central formado por bandas radiales de xilema que se extienden hacia el córtex externo entre las cuales se forman hileras de floema.
En las raíces aéreas, el cilindro de xilema, por lo general macizo en las raíces subterráneas, suele tener una médula central.
Desarrollo de la raíz
Las semillas de las plantas superiores contienen una planta que todavía no se desarrolla, esta es el embrión, cuando la semilla germina se observa la raíz embrionaria o radícula esta crece debido a la división de sus células y forma la Raíz primaria: Esta raíz tiene la función de posteriormente producir ramas. Raíz secundaria: Esta estructura consta de pelos absorbentes, tienen un sabor a corcho y es la parte principal de toda nuestra raíz.
Parénquima reservante. Acuífero y aerífero
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
El parénquima acuífero, está constituido por células de membranas sutiles, ricas en mucílagos, en las cuales redeposita el agua por inhibición del propio mucílago. Se presentan en algunas plantas de clima seco para mantener reservas de agua en el tallo y espesor de las hojas, (plantas suculentas), como, por ejemplo, las hojas de Aloe.
El parénquima aerífero es característico de plantas palustres y acuáticas donde sus células presentan amplios espacios intercelulares, facilitando el intercambio de gases en los órganos sumergidos.
En los amplios canales aeríferos como sucede en los pecíolos de las ninfeáceas, algunas células se transforman en idioblastos, situándose a lo largo de las filas de células de dichos canales. Efectúan estos una función mecánica pues dicho tejido es excesivamente blando. Las paredes de los ideoblastos presentan cristalitos de oxalato de calcio.
Las Plantas
La conservación de la ordenación específica que caracteriza a la vida, representa un trabajo que consume energía. La vida es, pues, un proceso ligado a la posibilidad de disponer de reservas energéticas suficientes para el mantenimiento del orden estructural. En el mismo momento en que cesa la aportación de energía, se extingue la vida activa y la ordenación comienza a desmoronarse.
El organismo animal obtiene la energía necesaria para la conservación de la vida exclusivamente del alimento, las plantas verdes, en cambio, la reciben inmediatamente de la luz solar. Entre animales y plantas verdes, existe, pues una diferencia fundamental en cuanto al modo de efectuar el aprovisionamiento de energía: las plantas, si se exceptúan algunos grupos y ciertas formas muy especializadas, son autótrofas, mientras que los animales heterótrofos. Todas las demás diferencias entre la organización animal y vegetal reposan, en último término, sobre esta diferencia básica.
Las plantas verdes toman de la luz solar la energía necesaria para la asimilación del carbono y de los demás elementos, no necesitan ser móviles como los animales, que deben ir en busca del alimento: las plantas típicas, de organización elevada, permanecen fijas, enraizadas en el substrato.
Existen, sin embargo, varias excepciones de esta regla. Hay, en efecto, grupos enteros de pequeños vegetales acuáticos, en su mayoría microscópicos, que durante toda su vida nadan libremente con la ayuda de órganos locomotores especiales.
Otra consecuencia del distinto modo de nutrirse animales y vegetales es la forma, fundamentalmente distinta, según la cual se produce el crecimiento en los dos grupos: tanto los animales como las plantas necesitan grandes superficies para la realización de los procesos metabólicos, pero mientras el desarrollo de los animales conduce, por medio de la precoz invaginación de una parte de la superficie, al establecimiento de amplias superficies internas, capaces de recibir y absorber el alimento, el de las plantas se orienta precisamente al revés, en el sentido de dar la mayor amplitud posible a las superficies externas asimiladoras, que puedan ser alcanzadas por la luz. Como los vegetales no necesitan cambiar de lugar, ningún límite externo se opone a su crecimiento; a diferencia de los animales, que siempre, después de un cierto período juvenil, cesan de crecer, las plantas continúan creciendo y desarrollando nuevos órganos hasta el momento de la su muerte. Se puede decir, pues, también, que el animal tiene una forma cerrada, mientras que la del vegetal es abierta.
También existen notables diferencias entre las células que forman a los vegetales y las que constituyen a los animales.
Las células vegetales
En el mundo viviente se encuentran básicamente dos tipos de células: las procarióticas y las eucarióticas. Las células procarióticas (del griego pro, antes de; karyon, núcleo) carecen de un núcleo bien definido. Todas las otras células del mundo animal y vegetal, contienen un núcleo rodeado por una doble membrana y se conocen como eucarióticas (del griego eu, verdadero y karyon, núcleo). En las células eucarióticas, el material genético DNA, está incluido en un núcleo distinto, rodeado por una membrana nuclear. Estas células presentan también varios organelos limitados por membranas que dividen el citoplasma celular en varios compartimientos, como son los cloroplastos, las mitocondrias, el retículo endoplasmático, el aparato de Golgi, vacuolas, etc.
Los organismos procariotes son unicelulares y pertenecen al grupo de las Moneras, que incluyen las bacterias y cianobacterias (algas verde-azules).
El DNA de las células procarióticas está confinado a una o más regiones nucleares, que se denominan nucleoides, que se encuentran rodeados por citoplasma, pero carecen de membrana.
En las bacterias, el núcleo está formado por un pedazo de DNA circular de aproximadamente 1 mm de largo, torcido en espiral, que constituye el material genético esencial. Las células procarióticas son las más primitivas de la tierra, hicieron su aparición en los océanos hace aproximadamente 3,5 millones de años; mientras que las células eucarióticas fósiles tienen menos de un millón de años.
Las células procarióticas son relativamente pequeñas, nunca tienen más de algunas micras de largo y no más de una micra de grosor. Las algas verde-azules son generalmente más grandes que las células bacterianas. Así mismo, todas las algas verde-azules realizan la fotosíntesis con la clorofila a, que no se encuentra en las bacterias, y mediante vías metabólicas comunes a las plantas y algas, pero no a las bacterias.
Un gran número de células procarióticas, están rodeadas por paredes celulares, que carecen de celulosa, lo que las hace diferentes de las paredes celulares de las plantas superiores.
En la parte interna de la pared celular, se encuentra la membrana plasmática o plasmalema, la cual puede ser lisa o puede tener invaginaciones, llamados mesosomas, donde se llevan a cabo las reacciones de transformación de energía (fotosíntesis y respiración). En el citoplasma, se encuentran cuerpos pequeños, esféricos, los ribosomas, donde se realiza la síntesis de proteínas. Así mismo, el citoplasma de las células procarióticas más complejas puede contener también vacuolas (estructuras en forma de saco), vesículas (pequeñas vacuolas) y depósitos de reserva de azucares complejos o materiales inorgánicos. En algunas algas verde-azules las vacuolas están llenas con nitrógeno gaseoso.
Muchas bacterias son capaces de moverse rápidamente gracias a la presencia de flagelos.
Células vegetales y animales
Tanto las células de las plantas como las de los animales son eucarióticas, sin embargo, presentan algunas diferencias:
Las células vegetales presentan una pared celular celulósica, rígida que evita cambios de forma y posición.
Las células vegetales contienen plastidios, estructuras rodeadas por una membrana, que sintetizan y almacenan alimentos. Los más comunes son los cloroplastos.
Casi todas las células vegetales poseen vacuolas, que tienen la función de transportar y almacenar nutrientes, agua y productos de desecho. Las células vegetales complejas, carecen de ciertos organelos, como los centriolos y los lisosomas.
Estructura primaria del tallo en dicotiledóneas
La estructura primaria es la que presentan los entrenudos del tallo mientras crecen en longitud. En esta estructura sólo existen tejidos primarios desarrollados a partir de los meristemas apicales (promeristemos),
Tejidos meristemáticos
Son tejidos embrionarios capaces de diferenciarse o perpetuarse; es decir, se multiplican activamente para formar los tejidos adultos diferenciados (crecimiento y especialización) y a su vez originan nuevas células meristemáticas. Los meristemas permiten que se produzca el crecimiento de las plantas en sentido longitudinal y diametral. El crecimiento longitudinal, también llamado crecimiento primario, se produce por la acción del meristema apical; mientras que el crecimiento diametral o en grosor, también denominado crecimiento secundario, se produce por divisiones que ocurren en el cambium vascular y, en menor proporción, en el cambium cortical.
Clasificación:
Según la posición topográfica en la planta:
Apicales: localizados en el ápice de los órganos (tallos, raíces, glándulas, etc.) que forman.
Basales: localizados en la base de los órganos (p.e., espinas) que forman.
Intercalares: situados entre células diferenciadas (maduras), contribuyendo al crecimiento en ambos lados.
Laterales: localizados en la periferia de los órganos, favoreciendo el crecimiento en grosor.
Axilares: son los meristemas apicales de las yemas, situados en las axilas de las hojas.
Según la naturaleza de las células que lo forman:
Primarios: provienen directamente de células que nunca han perdido su capacidad de división. (derivados de células embrionarias).
Secundarios: se originan a partir de células diferenciadas que nuevamente adquieren su capacidad de división.
Según el tiempo de aparición (secuencia de formación) en la planta:
Primarios: son los meristemas apicales del tallo y raíz presentes originariamente en el embrión y son capaces de producir los tejidos primarios.
Secundarios: se originan a partir de tejidos primarios por desdiferenciación. Producen los tejidos secundarios.
Características de las Células Meristemáticas.
Son células Indiferenciadas, pequeñas e isodiamétricas (excepto cámbium vascular). forman tejidos compactos, sin espacios intercelulares, gran núcleo (difuso) y poco citoplasma, pared celular delgada constituida de pared primaria y lamela media. (Algunos poseen campos de poros los), no poseen inclusiones citoplásmicas y con pocos orgánulos: abundantes ribosomas libres y dictiosomas, retículo endoplásmico (liso y rugoso) escaso. mitocondrias escasas y con pocas crestas, presentan proplastidios.
Meristemas Primarios (facilitan el crecimiento en longitud). Tipos.
Cuando ya la planta está formada, el tejido meristemático sólo se encuentra localizado en determinadas regiones del vegetal llamadas “zonas de crecimiento”, que conservan indefinidamente su carácter embrionario y su capacidad de división. Estas zonas se localizan en las yemas del tallo y en la región subterminal de las raíces. Los meristemos que aparecen en el extremo de los brotes o yemas y permiten el crecimiento en longitud de la planta, reciben el nombre de meristemos apicales.
Entre éstos se encuentran la protodermis, el procámbium, el meristema fundamental o básico.
Meristemas apicales: situados en los ápices de brotes y raíces (tanto principales como laterales), aquí se incluyen los meristemas caulinares, encargados del crecimiento del tallo, ramas y hojas y los meristemas apicales radicales, que son los responsables del crecimiento de las raíces.
Meristemas intercalares: situados en la base de los entrenudos de las ramas. (Aparecen más tarde en el tiempo).
Meristemas apicales caulinares:
Meristemas apicales radicales (yemas radicales):
Periciclo: Meristemo remanente propio de la raíz que se sitúa en la periferia del cilindro vascular. Colabora en la formación del córtex secundario, incluida la peridermis, del cambium vascular, y de las raíces laterales.
Meristemas Secundarios. Tipos. Se hacen cargo de facilitar el crecimiento en grosor.
El cambium vascular: Es un meristemo lateral formado por una monocapa cilíndrica de células situado en aquellos tallos y raíces que van a sufrir engrosamiento secundario. Sus células son de dos tipos: iniciales fusiformes e iniciales radiales, y se dividen en un plano tangencial al tallo o raíz. Las fusiformes dan lugar hacia el interior a células que se diferencian como xilema II y hacia el exterior a células que se diferencian como floema II. Las radiales dan lugar a parénquima. El cambium vascular del tallo se origina de dos formas: Bien a partir de células de procámbium presentes entre la xilema I y el floema I, dando lugar al cambium fascicular, bien por desdiferenciación de células parenquimáticas de los radios medulares, dando lugar al cambium interfascicular.
El cambium fascicular: es el cambium vascular que se forma a partir del procámbium situado entre la xilema y el floema primarios de los haces vasculares.
El cambium interfascicular: Es el cambium vascular que se forma por desdiferenciación de células del parénquima radiomedular, es decir, del parénquima situado entre los haces vasculares.
El cámbium suberógeno o felógeno: Meristemo lateral presente en los tallos y raíces de las plantas con engrosamiento secundario. Es una monocapa cilíndrica de células formada por la desdiferenciación de una capa de parénquima cortical subepidérmico. Por división tangencial estas células generan hacia el interior la felodermis y hacia el exterior el súber (corcho).
Las partes fundamentales que integran a esta estructura son: epidermis, corteza y estela o médula.
La epidermis, es el tejido más externo constituido por una capa delgada de células, se encuentra recubierto por una cutícula impermeable de cutina y, en la mayoría de los casos, pelos o tricomas.
Protodermis
Es una capa de células meristemáticas externa de los ápices vegetativo y radical que dará lugar a la epidermis.
La protodermis es una capa que se desarrolla sobre la superficie del ápice y sólo se divide en sentido anticlinal, es decir, perpendicular a la superficie. A través de la división de este meristema primario se origina la epidermis, la cual cumple las funciones de protección, contra agentes químicos y físicos externos y contra la desecación, además de permitir el intercambio gaseoso, secreción de compuestos metabólicos y absorción de agua en el cuerpo primario de la planta.
Meristemo fundamental o básico
Meristemo fundamental es un grupo de células meristemáticas que origina el resto de los tejidos de las plantas (parénquimas, colénquimas, esclerénquimas, etc.).
Procámbium
El Procámbium meristema primario constituido por una capa de células que originará los tejidos vasculares primarios (xilema y floema primarios). Puede formar, en algunos casos, el cámbium vascular.
Tejidos de protección. Epidermis
La epidermis procede de la capa más externa del promeristemo, la protodermis. Recubre externamente el cuerpo de la planta a modo de envoltura protectora, pero permite al mismo tiempo, el intercambio de materia con el medio exterior. Generalmente consta de una sola capa de células vivas, las que, en visión frontal, presentan aspecto poligonal o alargado y se disponen unidas unas a otras sin dejar espacios intercelulares constituyendo una película continua. A menudo el contorno lateral de las células es ondulado o dentado lo que refuerza la trabazón marginal de los elementos celulares.
La principal función de la epidermis es la de protección contra desecación y ataque de agentes externos, la misma tiene modificaciones como: los pelos o tricomas y estomas.
Tejidos de Protección. Modificaciones de la epidermis. Tricomas
Se da el nombre de tricoma, a todo apéndice epidérmico, independientemente de la forma que este tenga, que se encuentra a modo de resalto en la superficie de los órganos vegetales. Su forma, estructura y funciones, son diversos.
Las formas más comunes de tricomas son los pelos; también se consideran como tales a las papilas, las escamas y pelos absorbentes de las raíces. En cuanto a su estructura pueden ser unicelulares o pluricelulares, sencillos o ramificados y desempeñan diversas funciones: algunos son glandulares y producen sustancias volátiles que actúan como atractivos de insectos polinizadores o como repelentes de otros insectos, otros son urticantes (células de defensa) ya sea porque producen sustancias urticantes (hortiga) o porque tienen una constitución rígida y puntiaguda. Pueden encontrarse en todas las partes de la planta. Los tricomas deben estar formados exclusivamente por células epidérmicas
La corteza o córtex, está formada por todos los tejidos comprendidos entre la epidermis y el cilindro central, estela o médula, de la siguiente forma: la colénquima, ubicado inmediatamente después de la epidermis alternando con parénquima clorofílico;
Tejidos mecánicos o de sostén. Colénquima
Son los tejidos que le dan resistencia y elasticidad a la mayoría de las plantas.
Sin una cierta solidez la planta no podría conservar su forma.
Están constituidos por células dispuestas en una masa compacta y con las paredes celulares fuertemente engrosadas, bien en parte, como en la colénquima, o bien en su totalidad como en el esclerénquima.
Los principales tejidos mecánicos son: la colénquima y el esclerénquima.
La colénquima se caracteriza por ser un tejido vivo con protoplasto poco activo, sus células vistas en una sección longitudinal presentan una forma rectangular y transversalmente el aspecto más poligonal, la pared primaria es más gruesa en las esquinas (constituida por celulosa y sustancias pépticas), raras veces presentan cloroplastos, puede impregnarse de lignina y transformarse en esclerénquima.
La estructura y disposición de las células colenquimáticas en el cuerpo de la planta indican que la función primaria de este tejido es de sostén. Es el tejido típico de sostén, primero de los órganos en crecimiento y segundo de los órganos adultos herbáceos, presentándose característicamente en posición periférica en los tallos y hojas.
La estructura de la membrana o pared celular es el carácter más distintivo de las células colenquimáticas. Los espesamientos se disponen desigualmente, con cierta variabilidad en los distintos tipos de plantas. Una forma más común es presentar los espesamientos más importantes en los ángulos (colénquima angular), donde se reúnen varias células. Si el espesamiento no es tan manifiesto en los ángulos la cavidad celular adquiere una forma circular en su sección transversal (colénquima laminar). Son también conocidos como angular, laminar y lagunar.
El colénquima más especializado se origina conjuntamente con los tejidos vasculares a partir del procámbium y el menos especializado a partir del meristemo fundamental.
Tejidos parenquimáticos. Clorofílico
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas, presenta de las funciones metabólicas vacuola grande.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
El parénquima clorofílico es el parénquima que realiza una de las funciones más importantes del vegetal, la fijación del carbono utilizando las radiaciones solares. Ocupa las partes su epidérmicas de la planta hasta donde pueda penetrar la luz, posee células siempre vivas provistas de cloroplastos. Se encuentra más desarrollado en las hojas, donde forma una capa de tejido en empalizada y de parénquima lagunar.
Plastidios, Cloroplastos
Además del núcleo y las vacuolas, los cloroplastos, constituyen los organelos más conspicuos de una célula vegetal. Los cloroplastos están rodeados por una doble membrana, con una estructura interna constituida por un sistema de membranas, separadas por una matriz de naturaleza proteica llamada estroma, poseen DNA con una estructura similar al encontrado en células procarióticas, así como ribosomas, embebidos en el estroma. Todos los plastidios se desarrollan a partir de pro cloroplastos, que son cuerpos pequeños encontrados en plantas que crecen tanto en la luz como en la oscuridad. Se dividen por fisión o bipartición, como lo hacen las mitocondrias y los organismos procariotas. Los plastidios incoloros se conocen como leucoplastos, contienen enzimas responsables de la síntesis del almidón. Los leucoplastos mejor conocidos son los amiloplastos, que almacenan granos de almidón, como los encontrados en la raíz de la yuca, el tubérculo de la papa, en granos de cereales, etc. Otros leucoplastos pueden almacenar proteínas, se conocen como proteinoplastos. Los cromoplastos son organelos coloreados, especializados en sintetizar y almacenar pigmentos carotenoides (rojo, anaranjado y amarillo), estos son el origen de los colores de muchos frutos, flores y hojas, por ejemplo, la piel del tomate, la raíz de zanahoria, etc. Los cromoplastos se originan a partir de cloroplastos jóvenes o de cloroplastos maduros, por división.
Los cloroplastos son plastidios que contienen los pigmentos verdes clorofila a y b, así como carotenoides de color anaranjado y xantofilas amarillas, son característicos de los seres fotoautótrofos, que poseen la maquinaria enzimática para transformar la energía solar en energía química, a través de la fotosíntesis. Los cloroplastos son característicos de las células del mesófilo foliar, poseen una doble membrana que los asemeja a las mitocondrias. Tienen una membrana externa y otra interna, el espacio delimitado por la membrana interna está ocupado por un material amorfo, parecido a un gel, rico en enzimas, denominado estroma. Contiene las enzimas que realizan la fijación o reducción del CO2, convirtiéndolo en carbohidratos, como el almidón. La membrana interna de los cloroplastos también engloba un tercer sistema de membranas, que consta de sacos planos llamados tilacoides, en los cuales la energía luminosa se utiliza para oxidar el agua y formar ATP (compuesto rico en energía) y NADPH (poder reductor), usados en el estroma para convertir el CO2 en carbohidratos. En ciertas partes de los cloroplastos, los tilacoides se disponen como monedas apiladas, denominados grana, pero en el estroma permanecen aislados.
Los cloroplastos tienen forma elíptica, con un diámetro de 5 a 10 mm y su número puede variar de 20 a 100 por célula vegetal. Durante la ciclosis se mueven libremente en el citoplasma. Ellos responden directamente a la energía solar, para llevar a cabo la fotosíntesis, orientándose perpendicularmente a los rayos de luz ; sin embargo sí la energía lumínica es muy fuerte , se disponen de tal forma que la radiación incida oblicuamente, recibiendo menos luz.
Los cloroplastos se originan a partir de pro cloroplastos, reacción ésta que es disparada por la luz, que provoca la diferenciación del plastidios, apareciendo los pigmentos y la proliferación de membranas, que origina los tilacoides y grana. Así mismo, en el estroma del cloroplasto se encuentran pequeños pedazos circulares de DNA, dispuestos en doble hélice; parecidos al DNA de las mitocondrias y bacterias. El DNA del cloroplasto regula la síntesis del RNA ribosomal, del RNA de transferencia y de la Ribulosa 1,5 difosfato carboxilasa-oxigenasa (RUBISCO), enzima que cataliza la fijación del CO2 en la fotosíntesis. Sin embargo, la mayoría de las proteínas del cloroplasto, son sintetizadas en el citosol y transportadas al cloroplasto.
Las hojas
Las hojas son expansiones laterales de gran importancia funcional que se presentan en el tallo. Al conjunto de tallo y las hojas se les denomina brote, dado que no existe una separación clara entre ellos.
Hay una estrecha relación entre las hojas y el tallo pues la estructura de los tejidos conductores del tallo y las que se encuentran en el pecíolo y nervio principal de las hojas son semejantes.
Lo mismo puede decirse en cuanto a los tejidos parenquimáticos que se presentan en el córtex del tallo y el mesófilo de la hoja. No obstante, la hoja tiene unas funciones muy específicas: fotosíntesis y transpiración, para las cuales se encuentra muy especializada, tanto en su estructura como en su fisiología.
La forma laminar de la hoja, al aumentar la superficie de contacto con el medio, facilita la captación de energía luminosa y también sus intercambios gaseosos. Las hojas, a diferencia del tallo, no suelen presentar tejidos de reserva y están formadas fundamentalmente por tejidos primarios.
En las hojas llamadas sésiles, el limbo o lamina foliar se une directamente al tallo, mientras que en otras lo hace por medio de un pie llamado pecíolo, el cual se inserta en el tallo por la llamada base foliar.
En la mayoría de las monocotiledóneas y en unas pocas dicotiledóneas (Umbelíferas y Poligonáceas), la base de la hoja presenta un marcado ensanchamiento o vaina que rodea al tallo.
Las hojas son órganos vegetales con crecimiento limitado, normalmente con forma laminar, y estructura dorsiventral (tienen dos caras). Existe una gran diversidad de formas, colores, etc.
Estructura anatómica
La hoja posee una epidermis recubriendo toda la superficie externa. En su interior tiene un tejido parenquimático llamado mesófilo.
La epidermis está provista de estomas, y cubierta de una cutícula cérea cuyo grosor depende de las condiciones ambientales.
En el mesófilo de una hoja bifacial existen dos tipos de parénquima asimilador, organizado de la siguiente manera:
Las hojas Equifaciales son aquellas con exactamente la misma disposición en el haz y el envés, es decir, tiene los tejidos concéntricos unos a otros.
Existen estructuras anatómicas modificadas por las condiciones ambientales. Por ejemplo, en las plantas xerófitas, las hojas se encuentran reducidas debido a que las células están más concentradas para evitar la transpiración. La capa de ceras es más gruesa y una serie de pelos permiten mantener la humedad. Las hojas se pliegan sobre sí mismas, tapando las estomas.
En las hidrófitas, la epidermis prácticamente no tiene ceras. Los mesófilos tienen amplios espacios intercelulares donde se acumula el aire necesario para la respiración.
En cada hoja penetran uno, dos o tres haces vasculares diferentes, los cuales se dividen y constituyen la nervadura o nerviación de la hoja, y los haces del sistema conductor, xilema y floema. En los haces con mayor calibre hay fibras y colénquima, dando mayor consistencia a los órganos foliales. Los nervios principales están formados por un parénquima de transferencia amplio formado por varias capas de células. A esto lo suele recubrir una especie de vainas de células parenquimáticas que puede estar abierta o cerrada.
Morfología Foliar
En el exterior del tallo existen unas protuberancias sin diferenciación llamadas primordios foliares, teniendo crecimiento limitado. Normalmente el crecimiento de hojas se lleva a cabo por meristemas situados en posición apical y otros en posición basal. Y entre ellos pueden aparecer meristemos intercalares y laterales, aumentando la anchura de la hoja.
Habitualmente el meristemo apical cesa su crecimiento junto con el meristemo basal. Pero hay excepciones. existen dos tipos básicos de crecimiento de las hojas.
Además, los meristemos laterales ensanchan la hoja, en las dicotiledóneas esos meristemos laterales son subepidérmico, y pueden ocupar diferentes posiciones. en gramíneas y helechos los meristemos laterales son superficiales.
La hoja se divide en tres partes:
El limbo de las hojas habitualmente tiene simetría dorsiventral, y tiene dos caras, haz y envés o cara ventral y dorsal. El haz suele tener una coloración más oscura que el envés, y esto es resultado de la disposición de los tejidos. En cuánto a su Morfología, hay dos posibilidades:
Morfología del borde foliar: puede tener morfologías muy variadas. En muchos casos los entrantes y salientes se limitan a la periferia de la hoja, pero en otros casos, la hoja es lobulada. Si la incisión sobrepasa la mitad del limbo, la hoja es partida o secta. Dentro de este tipo hay varias divisiones, como las hojas pinnadas (la incisión se dirige hacia el nervio central), o palmadas
Las hojas de tipo compuesto son hojas partidas en las que cada corte llega al nervio central. Cada parte de la hoja se llama foliolo.
Si todos los foliolos parten del extremo del peciolo, sería una hoja palmado-compuesta. O puede ocurrir que los foliolos se inserten a los lados del nervio medio, y en este caso se llama pinnado-compuesta. En este caso el nervio central se llama raquis.
Nerviación de las hojas: se aprecian como unas costillas, más salientes por el envés de la hoja. En la hoja tiene, aparte de la función circulatoria, una función de soporte. Existen tres tipos básicos de nerviaciones:
La más primitiva es la dicótoma bifurcada o flabeliforme. Al final, los nervios llegan a contactar con el borde de la hoja. Por ello, también se le llama «abierta». Se encuentra en bastantes helechos y en un árbol llamado Ginkgo Biloba.
En los dos casos anteriores aparecen uno o varios nervios principales formando una red muy densa, y los nervios de menor tamaño contactan entre sí, y por ello se llaman «cerrados».
La nervadura reticulada (dicotiledóneas y helechos) tiene una o varias venas principales que se ramifican a su vez, pero cuando hay más de una vena principal siempre contactan en un punto. Existen varias modalidades, siendo la craspedódroma la más corriente. La nervadura actinódroma es parecida, pero en hojas palmadas. En la braquidódroma las nervaduras no llegan al borde y entran en contacto entre sí.
En la acródroma existen varias venas principales, las cuales discurren a lo largo de la hoja y se fusionan al llegar al final de la hoja. La campilódroma es parecida, pero más ensanchada y con la convergencia en el extremo.
En la nerviación paralela los nervios principales discurren paralelos unos a otros y siempre están separados en la base.
Existen dos modalidades: el tipo longitudinal estriado y arqueado estriado, y el tipo pinnado estriado.
En cuanto al peciolo, además de ser el punto por donde entran los haces conductores al limbo, actúa como soporte y mantiene al limbo estable. En unas ocasiones, es el tallo el que modifica su posición para exponer la hoja al sol, pero en otras ocasiones, el peciolo tiene un papel fundamental en el movimiento de la hoja. A veces, el peciolo tiene una almohadilla que se llama pulvímulo que actúa de articulación al hincharse o deshincharse. Incluso es posible que en la base de cada foliolo existan pulvímulos.
A veces el movimiento no se debe a la luz, sino que puede ser sensible a los roces, etc. Por ejemplo, la mimosa púdica, en cuanto se la toca, repliega totalmente los foliolos.
Normalmente, la base del peciolo tiene forma arqueada y con la parte abierta hacia el tallo.
Las hojas de una planta no son todas exactamente iguales, sino que varían su morfología según la edad y otros factores. En ocasiones, las hojas de la base son diferentes a las del ápice. A este fenómeno se le denomina anisofilia. Existe una modificación gradual. La heterofilia se da cuando existen en una misma planta hojas muy diferentes, y no hay cambio gradual.
Prefoliación: es la disposición de las hojas en el botón foliar. Previamente forma una especie de joroba que se va ensanchando, y finalmente adopta la forma definitiva de la hoja. La posición de la hoja puede variar de unas plantas a otras. Se representa con un punto la posición del eje del tallo.
Filotaxia o filotaxis es la disposición de las hojas a lo largo de un tallo adulto. En principio se entiende que existen tres tipos básicos:
En cuanto a la disposición verticilada, se cumplen dos premisas:
Los primordios foliares en la yema apical se encuentran en una disposición hexagonal, aunque en planos diferentes. Existen dos parámetros que determinan la posición de los primordios, el tamaño relativo de los primordios en relación al diámetro del cono apical del tallo y La posición relativa de los primordios foliares. Si forman un plano perpendicular al eje principal de la planta, se forman ortósticos.
Existen dos teorías que tratan de explicar la disposición final de las hojas dispersas:
Teoría clásica: es una teoría de naturaleza geométrica. Las hojas con este tipo de disposición se disponen a lo largo del tallo de tal forma que podemos trazar una hélice que une todos los puntos de inserción de las hojas, y en esa hélice vamos a poder situar todas las hojas. A esa hélice se le llama espiga fundamental o generatriz fundamental. La teoría establece que todas las hojas forman entre sí el mismo ángulo de divergencia.
Según esta teoría las hojas forman una serie matemática llamada serie de Fibonacci. Su primer término sería 1/2, el segundo 1/3, el tercero 2/5, 3/8, 5/13,… El numerador indica el número de vueltas que se da al tallo, y el denominador, el número de hojas que encontramos.
Se comprobó que los ángulos de divergencia entre los brotes se encontraban comprendidos entre 137º15´ y 138º. Es decir, el ángulo cambiaba en la zona joven o apical de la planta.
La serie de Fibonacci, como ángulo máximo tiene 137º30´29″. Por tanto, este modelo es bastante válido porque se aproxima a la realidad.
La teoría, sin embargo, tiene una serie de problemas:
La escuela francesa de botánica desarrolló la teoría de las hélices foliares múltiples.
No trata tanto de explicar de forma geométrica la situación, sino que explica el desarrollo de las hojas al crecer el tallo, y tiene muy en cuenta la anatomía de los ápices vegetativos.
Las hojas se insertan a lo largo del tallo distribuidas en series regulares que conforman un número variable de hélices. Va a haber tantas hélices como centros generadores de hojas existan en el ápice del tallo. También sustituye el concepto de ángulo de divergencia por el de contigüidad. Establecen tres tipos diferentes de contigüidad:
Esto se encuentra en consonancia con la disposición de los primordios foliares en el ápice vegetativo del tallo.
Polimorfismo foliar y adaptaciones especiales
La forma de las hojas no sólo depende de la especie, sino que presenta variaciones a lo largo de la misma planta. Las hojas embrionarias son los cotiledones, hojas ya presentes en la semilla, carnosas y repletas de sustancias de reserva. A continuación, se encuentran los catafilos, que aparecen en todas las plantas, de aspecto escamoso y sin clorofila, y se encuentran entre los cotiledones y las primeras hojas de aspecto normal. También se les llama catafilos a las escamas que recubren las yemas enervantes, también llamadas a veces pérulas o tegumentos.
También se encuentran hojas tectrices, en cuya axila aparece una yema lateral. Otro tipo son los profilos, las primeras hojas que se forman cuando empieza a crecer una yema lateral. También hay hipsofilos, hojas de tamaño más reducido que aparecen cerca de las inflorescencias, de aspecto más sencillo que el de las hojas normales de la planta.
Las brácteas son hojas modificadas que se encuentran muy cerca de las flores. La modificación es morfológica, aunque a veces también varía el color. Las flores pueden considerarse hojas modificadas. Se llamarían tépalos a las piezas de la envuelta floral cuando no hay diferencia entre cáliz y corola. A su conjunto se le llama perigonio. Los pétalos son cada una de las piezas corola, y los sépalos cada una de las piezas del cáliz.
Los estambres son hojas modificadas, cuyo conjunto constituye hojas modificadas. El gineceo está formado por carpelos, y también son hojas muy modificadas.
En cuanto a las adaptaciones especiales, se conocen casos de hojas espinosas, como en el acebo y en la encina. Cuando se trata de hojas compuestas, es más complicado, transformándose en espinas la estípula, el raquis, etc.
Hay muchas hojas que se transforman en zarcillos, total o parcialmente. Se encuentran en muchas leguminosas.
Otra modificación son los filodios, hojas con nerviación paralela, y que en realidad corresponden a un peciolo aplanado, como muchas especies de acacia.
También es frecuente encontrar hojas transformadas en órganos de reserva, como cebollas y otras plantas que tienen bulbos basales. También se consideran órganos de reserva a las hojas carnosas de las Crasuláceas.
Las hojas xeromórfica son de pequeño tamaño. Además, tienen una consistencia coriácea. Siempre tienen una gruesa cutícula, y suelen tener pelos entre las estomas hundidos. También repliegan su limbo para encerrar las estomas en su interior. Es muy frecuente que sean Equifaciales.
Las hojas de las plantas insectívoras son de plantas que viven en ambientes muy pobres que se adaptan a capturar pequeños animales, por lo que es indispensable la modificación para captarlos. Ejemplos:
Tejidos parenquimáticos. Clorofílico en empalizada
El parénquima de la hoja vegetativa, el mesófilo, está limitado en sus dos caras por una epidermis. La diferenciación anatómica de dicho parénquima está en correspondencia, con las funciones principales de las hojas, la fotosíntesis de los hidratos de carbono y el desprendimiento de vapor de agua. El parénquima asimilador, rico en cloroplastos, dispuesto bajo la epidermis de la cara superior, se conoce en la mayoría de las hojas de una o varias capas de células cilíndricas dispuestas perpendicularmente a la superficie y recibe a causa de su aspecto en la sección transversal de la hoja, el nombre de parénquima en empalizada; en éste, las células que lo forman, están muy unidas entre sí y son ricas en cloroplastos.
En las hojas con estructura isolateral, el parénquima clorofílico en empalizada se encuentra presente en ambas caras de las hojas.
parénquima cortical, formado por varias células, de forma, tamaño y disposición irregular, guarda dentro de sí varias cámaras intercelulares; también células secretoras llamadas epitélicas, a pesar de ser raras, se presentan en forma abundante en algunas plantas; y la vaina amilífera que es la que limita la corteza del cilindro central o estela.
Periciclo: Meristemo remanente propio de la raíz que se sitúa en la periferia del cilindro vascular. Colabora en la formación del córtex secundario, incluida la peridermis, del cambium vascular, y de las raíces laterales.
Meristemas Secundarios. Tipos. Se hacen cargo de facilitar el crecimiento en grosor.
El cambium vascular: Es un meristemo lateral formado por una monocapa cilíndrica de células situado en aquellos tallos y raíces que van a sufrir engrosamiento secundario. Sus células son de dos tipos: iniciales fusiformes e iniciales radiales, y se dividen en un plano tangencial al tallo o raíz. Las fusiformes dan lugar hacia el interior a células que se diferencian como xilema II y hacia el exterior a células que se diferencian como floema II. Las radiales dan lugar a parénquima. El cambium vascular del tallo se origina de dos formas: Bien a partir de células de procámbium presentes entre el xilema I y el floema I, dando lugar al cambium fascicular, bien por desdiferenciación de células parenquimáticas de los radios medulares, dando lugar al cambium interfascicular.
El cambium fascicular: es el cambium vascular que se forma a partir del procámbium situado entre la xilema y el floema primarios de los haces vasculares.
El cambium interfascicular: Es el cambium vascular que se forma por desdiferenciación de células del parénquima radiomedular, es decir, del parénquima situado entre los haces vasculares .
El cámbium suberógeno o felógeno: Meristemo lateral presente en los tallos y raíces de las plantas con engrosamiento secundario. Es una monocapa cilíndrica de células formada por la desdiferenciación de una capa de parénquima cortical subepidérmico. Por división tangencial estas células generan hacia el interior la felodermis y hacia el exterior el súber (corcho).
| La estela o cilindro central, es la parte central del cuerpo primario del tallo, está formada por el periciclo, los tejidos vasculares, los radios medulares y la médula. El periciclo, se encuentra formando casquete encima del floema. Tejidos Vasculares Primarios. En el tallo primario, la xilema y el floema se agrupan para formar los haces vasculares, que al microscopio se observan como estructuras ovaladas y se les denomina haces colaterales abiertos. |
Los radios medulares: están formados por tejidos parenquimáticos, que se disponen entre los haces vasculares y que comunican la corteza con la médula.
La médula, no es muy común encontrarla en tallos de plantas monocotiledóneas, es la región central y está constituida por tejido parenquimático de almacenamiento o de relleno.
Tejidos vasculares o conductores. Xilema
Es el encargado de transportar de sustancias no elaboradas (savia bruta) a través de todo el cuerpo de las plantas traqueophytas. En las plantas que no poseen tejidos vasculares como las BRIOPHYTAS (musgos), Talophytas (algas, hongos y bacterias) no sucede este proceso.
El xilema es el principal tejido conductor de agua y de sustancias inorgánicas (minerales) y el floema es el tejido conductor de las sustancias alimenticias de una parte de la planta a otra. O sea, los principales tejidos componentes del sistema vascular están representados por la xilema y el floema.
La importancia fisiológica y filogenético del sistema vascular y su destacado papel entre los elementos estructurales del cuerpo de la planta determinó la segregación taxonómica de las plantas provistas de dicho sistema, formando el grupo de las llamadas plantas vasculares.
Xilema:
Al estudiar los meristemos primarios observamos, que el procámbium se localiza dentro del embrión como una banda diferenciada que rompe la continuidad del meristema fundamental o básico, el origina corteza y médula.
El xilema o leño que se diferencia en el cuerpo primario de la planta, se denomina xilema primaria.
Si la planta es de tal naturaleza que después de terminar el crecimiento primario forma tejidos secundarios mediante la actividad del cambium vascular, la xilema formado por este meristemo constituye la xilema secundaria.
El tejido más interno de la estela o cilindro central, adyacente a la médula si la misma está presente, es la xilema primaria.
Este es un tejido compuesto por los siguientes por los siguientes elementos:
Traqueidas
Son las células vasculares donde cada una de ellas queda limitada por membranas transversales primitivas conservando, por lo tanto, la integridad inicial de sus membranas por lo que se les llama vasos cerrados.
Estos vasos son simples células muertas, alargadas y generalmente aguzadas en los extremos, las paredes son delgadas en ciertas regiones alcanzando considerable espesor en otras, producido por la presencia de engrosamientos locales internos.
En la mayoría de las Gimnospermas y algunas Angiospermas menos evolucionadas las traqueidas son los únicos elementos conductores de agua y además sirven como elementos de sostén.
Tráqueas
Son células vasculares donde el enlace de una fila de células superpuestas se produce por resorción de las membranas divisorias primitivas (por lo que también se llaman vasos abiertos).
Por lo antes expuesto, entendemos que una tráquea no es una sola célula, sino un tubo formado por una serie de células cilíndricas muertas, las mayorías de las cuales han disuelto las paredes de los extremos. Estos tubos en la xilema primaria, como las traqueidas, presentan engrosamientos locales internos que pueden ser que pueden ser de los siguientes tipos: anular, espiralado, escalariforme y punteado.
Las tráqueas tienen la pared mucho más engrosada y generalmente lignificada.
Haciendo una comparación entre las traqueidas y las tráqueas vemos que difieren en que las traqueidas poseen células imperforadas, únicamente provistas de pares de puntuaciones en sus membranas, mientras que las membranas de las células que forman las tráqueas están perforadas en ciertas áreas de contacto con otras células. De este modo los elementos de los vasos se unen unos con otros, formando largos tubos continuos, por ello, las traqueidas son células aisladas tubulosas y las tráqueas son tubos relativamente anchos, amplios más raramente estrechos.
Fibras de la xilema primaria (células esclerenquimáticas).
Las fibras de la xilema primaria se derivan del procámbium. Son células alargadas con paredes engrosadas totalmente y por lo general, puntiagudas en los extremos; en algunos casos tienen hasta 1m de largo, pero no es lo común. En ocasiones el engrosamiento de la pared celular es tan grande que la cavidad (lumen) de la célula casi desaparece.
Cuando las fibras han alcanzado su longitud completa y el máximo del engrosamiento de la pared, los protoplastos mueren. Antes de que esto ocurra las paredes se impregnan de lignina.
Esta modificación química de las paredes produce un aumento de la fuerza y dureza del vegetal.
Frecuentemente, las fibras son muy elásticas y se pueden estirar mucho sin perder su capacidad de recuperar su tamaño original.
Células parénquimaticas xilemáticas:
Las células del parénquima están mezcladas con los otros elementos componentes de la xilema primaria. Estas células forman radios medulares por los filos verticales de células vivas en el xilema.
El parénquima asociado con los tejidos vasculares primarios se origina en el procambium.
Las células parénquimaticas xilemáticas pueden ser largas, con contenido variado de sustancias como almidón, grasas, taninos, cristales y otros.
En sus paredes únicamente se encuentran pares de puntuaciones simples.
Xilema secundario
La xilema secundaria también está constituida por varios elementos:
Traqueidas, tráqueas, células parénquimaticas, radios medulares y en ocasiones células secretoras.
Por lo general aparece en las Gimnospermas y en las plantas Dicotiledóneas con crecimiento secundario, y se origina por divisiones tangenciales de las células del cambium vascular, las que se encuentran ordenadas paralelamente a los radios del cuerpo secundario de la planta.
El leño, como también se le denomina, presenta generalmente sus células lignificadas, o sea, con membranas duras y rígidas; en las Dicotiledóneas se componen comúnmente de tres clases distintas de tejidos, con membranas más o menos lignificadas:
Los vasos están representados por traqueidas y tráqueas, las que sirven para el transporte de agua y le proporcionan gran resistencia al tallo. Las traqueidas están formadas por elementos muy amplios y cortos. Estos vasos son del tipo escalariforme o punteado.
Se considera que en las Dicotiledóneas las Traqueidas y las fibras llegan a ser considerablemente más largas que las células de las cuales derivan.
Las fibras leñosas son, por lo general, muy estrechas y largas, con los extremos aguzados y con las membranas gruesas provistas de escasas punteaduras. En dichas fibras el contenido protoplasmático ha sido reabsorbido.
Tílides
En las partes más profundas del leño de muchas plantas, por ejemplo, de Ficus, Vitis, Quercus, se pueden observar pequeñas hernias en el interior de los vasos leñosos que los obstruyen en mayor o en menor grado, los cuales se le denomina tílides. Son formados por abolladuras de las membranas de células parenquimáticas que se hallan siempre circundando los vasos en forma de anillo y que pueden vaciarse completamente o transformarse en elementos reservantes, acumuladores de almidón. Se desarrollan a través de los pares de puntuaciones que relacionan las células parenquimáticas con los elementos traqueales.
Tejidos vasculares o conductores. Floema
Mientras que la xilema es el principal tejido conductor de agua y de sustancias inorgánicas (minerales), el floema es el tejido conductor de las sustancias alimenticias de una parte de la planta a otra.
El segundo tejido de la estela o cilindro central, originado por el procámbium hacia el exterior de la xilema primaria, es el floema primario.
Floema primario. Consta de varios elementos componentes:
Tubos o vasos cribosos:
Sirven para el transporte de las materias orgánicas elaboradas por las plantas, a partir del proceso de fotosíntesis, se componen de células vivientes, dispuestas en series longitudinales y unidos entre sí por medio de poros abiertos que resultan de la perforación local de la membrana divisoria entre dos células procambiales contiguas. Las células de los tubos cribosos presentan protoplastos vivos, muy pobres en plasma. Es un hecho notable que sus núcleos desaparecen muy pronto; presentan leucoplastos, en los que a menudo se observa almidón y, como jugo celular poseen un líquido acuoso que durante el período de asimilación es especialmente rico en sacarosa y a veces también en prótidos.
En general los tubos cribosos son activos únicamente durante un solo período de vegetación. Hacia el final de dicho período, las cribas quedan obstruidas por gruesas acumulaciones de calos, sustancias hialinas que ya mucho antes revestían de una débil película las paredes de los poros. Debido a ello queda interrumpido el intercambio de materia entre los elementos de los tubos cribosos.
Células anexas o acompañantes:
Adyacentes a los tubos cribosos, existen una o más hileras paralelas de células asociadas, que pueden ser de la misma longitud o más cortas que las células que componen los tubos cribosos.
La célula inicial de las anexas tiene el lumen mucho más estrecho que los tubos cribosos, son muy ricas en plasmas, poseen grandes núcleos, a través de un gran número de finas punteaduras y plasmodesmos se comunican con los tubos cribosos y asumen el control de las actividades que ocurren en el plasma de éstos.
Células parénquimaticas:
El floema contiene, además de las células acompañantes, células parenquimáticas, en cantidad variable, y son las encargadas del almacenamiento de grasa, almidón y otros materiales orgánicos alimenticios y acumulaciones de taninos y resinas.
Estas células son alargadas y están orientadas de forma que sus ejes longitudinales son paralelos a la dirección longitudinal del tejido vascular.
Fibras:
Pueden alcanzar gran longitud. Sus células forman membranas secundarias después de complementar su alargamiento. En algunas plantas las fibras están lignificadas típicamente, en otras no. Las puntuaciones son por lo general simples; también pueden presentarse rebordeadas o areoladas.
Tejidos vasculares de origen secundario:
En la mayoría de las Dicotiledóneas y Gimnospermas, los tejidos vasculares primarios en el tallo están dispuestos en forma de haces agrupados circularmente; con la xilema hacia el interior y el floema hacia el exterior. Como ya se ha dicho, una parte de las células del procámbium permanecen meristemáticas aún después de completarse el crecimiento primario, desarrollándose por consiguiente el cambium vascular en forma de un cilindro ininterrumpido entre los haces vasculares vecinos.
Este cilindro discontinuo pronto se hace continuo ya que las capas de parénquima entre los haces cercanos manifiestan actividad meristemática, formando el cambium interfascicular. De esta manera, el cambium vascular en el tallo está constituido de porciones alternas de cambium fascicular e interfascicular.
El cambium fascicular por repetidas divisiones mitóticas de sus células forma leño hacia la parte interna y liber hacia la parte externa, mientras que el cambium interfascicular forma además parénquima radiomedular. En algunas Dicotiledóneas herbáceas con crecimiento secundario, el cambium interfascicular produce solamente fibras o sólo parénquima esclerotizado en el lado de la xilema (Medicago), o bien el crecimiento secundario puede ser tan pequeño que queda limitado a los haces vasculares (Cucúrbita).
En la raíz de Dicotiledóneas y Gimnospermas, la parte principal del cambium vascular, se forma entre la xilema primaria y el floema en la estela. Cuando se produce crecimiento secundario en ella, una capa de células de procámbium se forma entre la xilema y el floema, se conserva permanente embriónico y no diferenciado, representando una parte del cambium vascular de la raíz. El resto forma poco después las células parénquimaticas del periciclo. Este cambium vascular se presenta primero el borde interno de los cordones floemáticas.
Floema secundario
El líber secundario se forma por la actividad del cambium hacia el interior de los cordones liberianos primarios y constituyen la parte principal de la corteza secundaria; está formado por diversos elementos que se superponen de manera regular. Estos elementos son:
Los tubos cribosos, con placas perforadas, y en las Dicotiledóneas acompañadas de células anexas ricas en plasma.
El parénquima liberiano, constituido de células de membranas sutiles, que se llenan de almidón o de sustancias proteicas, (como en las Gimnospermas).
Las fibras esclerenquimáticas (por lo general muertas).
Se puede observar, además, esclereidas, radios medulares, células crístaliferas, secretoras, tubos laticíferos y células mucilaginíferas.
En ciertas Dicotiledóneas se presenta una gran diversidad de tipos de ordenación en sus células, presentándose las fibras dispuestas en bandas tangenciales, alternando más o menos regularmente con bandas de tubos cribosos, que por lo general forman conductos longitudinales ininterrumpidos y ramificados que van desde las raíces a las hojas.
Estos tubos limitan al que los cordones de parénquima liberiano, con los radios medulares, también longitudinales, los que constituyen la prolongación radical de los radios medulares del leño.
Tejidos vasculares o conductores. Haces
Una porción del sistema vascular primario del tallo o de la hoja, constituye un cordón o haz vascular.
El floema y la xilema se hallan asociados no sólo en su conjunto, sino también por lo regular dentro de sus partes, es decir, en los haces vasculares. Las diferentes maneras de disponerse los tejidos vasculares dentro de los haces ha permitido clasificarlos en distintos tipos:
Haces colaterales abiertos:
Son aquellos que conservan algunas células meristemáticas (o procambiales) entre el floema (líber) y el xilema (leño), productoras de elementos secundarios con origen primario.
Estas células procambiales constituyen una zona generatriz, integrada por células meristemáticas situadas entre el floema y la xilema, la cual por repetidas divisiones tangenciales de sus células produce xilema secundaria hacia la parte interna y floema secundario hacia el exterior. Poseen cambium en general las Gimnospermas y las Dicotiledóneas y su actividad se debe al crecimiento secundario de los mismos. El cambium que se halla entre la xilema y el floema se llama cambium fascicular.
Haces colaterales cerrados:
Se consideran cerrados los haces donde el floema y la xilema están en contacto inmediato, incapaces de modificar su estructura primitiva. Este tipo de haz es característico de las plantas monocotiledóneas. En estos tipos de haces todas las células del procámbium se diferencian en elementos de la xilema y el floema y ninguno permanece en estado meristemático como aquellos del cambium fascicular de las Gimnospermas y Dicotiledóneas. Debido a la ausencia del cambium, la mayoría de las monocotiledóneas no experimentan crecimiento en grosor.
En los haces conductores existen generalmente vainas de tejidos mecánicos en el lado exterior del floema y, a veces también en lado interno del haz, por fuera de la xilema; estas vainas en sección transversal aparecen como extractos semilunares de esclerénquima. Son pues, incompletos en los haces abiertos. A cada lado del haz, es decir, junto al límite entre la xilema y el floema, la vaina se convierte en una banda de células parenquimáticas más débilmente engrosadas, que permiten el intercambio de agua y materias alimenticias entre el haz y el parénquima circundante.
Haces bicolaterales:
Se presentan en determinadas familias de plantas como Cucurbitáceas y Solanáceas; estas poseen dos zonas de floema: una en posición normal, como en el haz colateral y la otra más hacia adentro que el leño y menos desarrollado que la primera.
Haces concéntricos:
En algunas Pteridofitas y Monocotiledóneas el tipo de haz es denominado concéntrico; los hay de dos clases atendiendo a la posición de los tejidos vasculares. Con la xilema en la parte externa rodeando el floema, (tipo frecuente en los rizomas de las Monocotiledóneas), llamándose por ello concéntrico perixilemático. Y con la xilema en la parte central y el floema al exterior (frecuentemente se presenta en los helechos, por ejemplo, en el género Pteris), se designa con el nombre de concéntrico perifloemático.
Haces radiales:
Se encuentra en la raíz y es característico de este órgano. Este haz conductor radial comprende varios cordones xilemáticas y floemáticas, que se disponen como los radios de una rueda, alternando los de una clase con lo de la otra. El conjunto de los cordones xilemáticos de ordinario aplanado lateralmente, presenta pues forma estrellada en la sección transversal. Los entrantes comprendidos entre las láminas leñosas están ocupados por los cordones cribosos, que quedan separados de los cordones xilemáticos por una o varias capas de células parenquimáticas.
En los haces radiales, la formación de todos los elementos de los cordones xilemáticos y floemáticos avanzan progresivamente de la periferia del haz hacia el centro del mismo, en la porción más externa quedan por ello las células primiciales o primarias (el protoxilema y protofloema); les sigue hacia el interior los elementos diferenciados más tarde (metaxilema y metafloema). Estos haces son típicos de raíces; en el vástago se presenta raramente, por ejemplo, en algunos helechos. El número de radios xilemáticos varía de unas especies a otras.
El microscópio
Microscopio, cualquiera de los distintos tipos de instrumentos que se utilizan para obtener una imagen aumentada de objetos minúsculos o detalles muy pequeños de los mismos.
Microscopio óptico
El tipo de microscopio más utilizado es el microscopio óptico, que se sirve de la luz visible para crear una imagen aumentada del objeto. El microscopio óptico más simple es la lente convexa doble con una distancia focal corta. Estas lentes pueden aumentar un objeto hasta 15 veces. Por lo general se utilizan microscopios compuestos, que disponen de varias lentes con las que se consiguen aumentos mayores. Algunos microscopios ópticos pueden aumentar un objeto por encima de las 2.000 veces.
El microscopio compuesto consiste en dos sistemas de lentes, el objetivo y el ocular, montados en extremos opuestos de un tubo cerrado. El objetivo está compuesto de varias lentes que crean una imagen real aumentada del objeto examinado. Las lentes de los microscopios están dispuestas de forma que el objetivo se encuentre en el punto focal del ocular. Cuando se mira a través del ocular se ve una imagen virtual aumentada de la imagen real. El aumento total del microscopio depende de las longitudes focales de los dos sistemas de lentes.
El equipamiento adicional de un microscopio consta de un armazón con un soporte que sostiene el material examinado y de un mecanismo que permite acercar y alejar el tubo para enfocar la muestra. Los especímenes o muestras que se examinan con un microscopio son transparentes y se observan con una luz que los atraviesa, y se suelen colocar sobre un rectángulo fino de vidrio. El soporte tiene un orificio por el que pasa la luz. Bajo el soporte se encuentra un espejo que refleja la luz para que atraviese el espécimen. El microscopio puede contar con una fuente de luz eléctrica que dirige la luz a través de la muestra.
La fotomicrografía, que consiste en fotografiar objetos a través de un microscopio, utiliza una cámara montada por encima del ocular del microscopio. La cámara suele carecer de objetivo, ya que el microscopio actúa como tal. El término microfotografía, utilizado a veces en lugar de fotomicrografía, se refiere a una técnica de duplicación y reducción de fotografías y documentos a un tamaño minúsculo para guardarlos en un archivo.
Los microscopios que se utilizan en entornos científicos cuentan con varias mejoras que permiten un estudio integral del espécimen. Dado que la imagen de la muestra está ampliada muchas veces e invertida, es difícil moverla de forma manual. Por ello los soportes de los microscopios científicos de alta potencia están montados en una plataforma que puede moverse con tornillos micrométricos. Algunos microscopios cuentan con soportes giratorios. Todos los microscopios de investigación cuentan con tres o más objetivos montados en un cabezal móvil que permite variar la potencia de aumento.
Microscopios ópticos especiales
Hay diversos microscopios ópticos para funciones especiales. Uno de ellos es el microscopio estereoscópico, que no es sino un par de microscopios de baja potencia colocados de forma que convergen en el espécimen. Estos instrumentos producen una imagen tridimensional.
El microscopio de luz ultravioleta utiliza el rango ultravioleta del espectro luminoso en lugar del rango visible, bien para aumentar la resolución con una longitud de onda menor o para mejorar el detalle absorbiendo selectivamente distintas longitudes de onda de la banda ultravioleta. Dado que el vidrio no transmite las longitudes de onda más cortas de la luz ultravioleta, los elementos ópticos de estos microscopios están hechos con cuarzo, fluorita o sistemas de espejos aluminizados. Además, dado que la radiación ultravioleta es invisible, la imagen se muestra con fosforescencia (véase Luminiscencia), en fotografía o con un escáner electrónico. El microscopio de luz ultravioleta se utiliza en la investigación científica.
El microscopio petrográfico o de polarización se utiliza para identificar y estimar cuantitativamente los componentes minerales de las rocas ígneas y las rocas metamórficas. Cuenta con un prisma de Nicol u otro tipo de dispositivo para polarizar la luz que pasa a través del espécimen examinado (véase Óptica: Polarización de la luz). Otro prisma Nicol o analizador determina la polarización de la luz que ha pasado a través del espécimen. El microscopio tiene un soporte giratorio que indica el cambio de polarización acusado por el espécimen.
El microscopio en campo oscuro utiliza una luz muy intensa en forma de un cono hueco concentrado sobre el espécimen. El campo de visión del objetivo se encuentra en la zona hueca del cono de luz y sólo recoge la luz que se refleja en el objeto. Por ello las porciones claras del espécimen aparecen como un fondo oscuro y los objetos minúsculos que se están analizando aparecen como una luz brillante sobre el fondo. Esta forma de iluminación se utiliza para analizar elementos biológicos transparentes y sin manchas, invisibles con iluminación normal.
El microscopio de fase ilumina el espécimen con un cono hueco de luz, como en el microscopio en campo oscuro. Sin embargo, en el microscopio de fase el cono de luz es más estrecho y entra en el campo de visión del objetivo, que contiene un dispositivo en forma de anillo que reduce la intensidad de la luz y provoca un cambio de fase de un cuarto de la longitud de onda. Este tipo de iluminación provoca variaciones minúsculas en el índice de refracción de un espécimen transparente, haciéndolo visible. Este tipo de microscopio es muy útil a la hora de examinar tejidos vivos, por lo que se utiliza con frecuencia en biología y medicina.
Entre los microscopios avanzados se encuentran el microscopio de campo cercano, con el que pueden verse detalles algo menores a la longitud de onda de la luz. Se hace pasar un haz de luz a través de un orificio diminuto y se proyecta a través del espécimen a una distancia equivalente a la mitad del diámetro del orificio, formando una imagen completa.
Microscópio electrónico
La potencia amplificadora de un microscopio óptico está limitada por la longitud de onda de la luz visible. El microscopio electrónico utiliza electrones para iluminar un objeto. Dado que los electrones tienen una longitud de onda mucho menor que la de la luz pueden mostrar estructuras mucho más pequeñas. La longitud de onda más corta de la luz visible es de alrededor de 4.000 Ángstrom (1 ángstrom es 0,0000000001 metros). La longitud de onda de los electrones que se utilizan en los microscopios electrónicos es de alrededor de 0,5 ángstroms.
Todos los microscopios electrónicos cuentan con varios elementos básicos. Disponen de un cañón de electrones que emite los electrones que chocan contra el espécimen, creando una imagen aumentada. Se utilizan lentes magnéticas para crear campos que dirigen y enfocan el haz de electrones, ya que las lentes convencionales utilizadas en los microscopios ópticos no funcionan con los electrones. El sistema de vacío es una parte relevante del microscopio electrónico. Los electrones pueden ser desviados por las moléculas del aire, de forma que tiene que hacerse un vacío casi total en el interior de un microscopio de estas características. Por último, todos los microscopios electrónicos cuentan con un sistema que registra o muestra la imagen que producen los electrones.
Hay dos tipos básicos de microscopios electrónicos: el microscopio electrónico de transmisión (Transmission Electron Microscope, TEM) y el microscopio electrónico de barrido (Scanning Electron Microscope, SEM). Un TEM dirige el haz de electrones hacia el objeto que se desea aumentar. Una parte de los electrones rebotan o son absorbidos por el objeto y otros lo atraviesan formando una imagen aumentada del espécimen. Para utilizar un TEM debe cortarse la muestra en capas finas, no mayores de un par de miles de ángstroms. Se coloca una placa fotográfica o una pantalla fluorescente detrás del objeto para registrar la imagen aumentada. Los microscopios electrónicos de transmisión pueden aumentar un objeto hasta un millón de veces.
Un microscopio electrónico de barrido crea una imagen ampliada de la superficie de un objeto. No es necesario cortar el objeto en capas para observarlo con un SEM, sino que puede colocarse en el microscopio con muy pocos preparativos. El SEM explora la superficie de la imagen punto por punto, al contrario que el TEM, que examina una gran parte de la muestra cada vez. Su funcionamiento se basa en recorrer la muestra con un haz muy concentrado de electrones, de forma parecida al barrido de un haz de electrones por la pantalla de una televisión. Los electrones del haz pueden dispersarse de la muestra o provocar la aparición de electrones secundarios. Los electrones perdidos y los secundarios son recogidos y contados por un dispositivo electrónico situado a los lados del espécimen. Cada punto leído de la muestra corresponde a un píxel en un monitor de televisión. Cuanto mayor sea el número de electrones contados por el dispositivo, mayor será el brillo del píxel en la pantalla. A medida que el haz de electrones barre la muestra, se presenta toda la imagen de la misma en el monitor. Los microscopios electrónicos de barrido pueden ampliar los objetos 100.000 veces o más. Este tipo de microscopio es muy útil porque, al contrario que los TEM o los microscopios ópticos, produce imágenes tridimensionales realistas de la superficie del objeto.
Se han desarrollado otros tipos de microscopios electrónicos. Un microscopio electrónico de barrido y transmisión (Scanning Trasnmission Electron Microscope, STEM) combina los elementos de un SEM y un TEM, y puede mostrar los átomos individuales de un objeto. El microanalizador de sonda de electrones, un microscopio electrónico que cuenta con un analizador de espectro de rayos X, puede analizar los rayos X de alta energía que produce el objeto al ser bombardeado con electrones. Dado que la identidad de los diferentes átomos y moléculas de un material se puede conocer utilizando sus emisiones de rayos X, los analizadores de sonda de electrones no sólo proporcionan una imagen ampliada de la muestra, como hace un microscopio electrónico, sino que suministra también información sobre la composición química del material.
Microscópio de sonda de barrido
En los microscopios de sonda de barrido se utiliza una sonda que recorre la superficie de una muestra, proporcionando una imagen tridimensional de la red de átomos o moléculas que la componen. La sonda es una afilada punta de metal que puede tener un grosor de un solo átomo en su extremo. Un tipo importante de microscopio de sonda de barrido es el microscopio de túnel de barrido (siglas en inglés de Scanning Tunelling Microscope, STM) desarrollado en 1981. Este microscopio utiliza un fenómeno de la física cuántica, denominado efecto túnel, para proporcionar imágenes detalladas de sustancias conductoras de electricidad. La sonda se coloca a una distancia de pocos ángstroms de la superficie del material y se aplica un voltaje pequeño entre la superficie y la sonda. A causa de la poca distancia entre el material y la sonda algunos electrones se escapan a través del hueco, generando una corriente. La magnitud de la corriente del efecto túnel depende de la distancia entre la superficie y la sonda. El flujo de corriente es mayor cuando la sonda se acerca al material y disminuye cuando se aleja. A medida que el mecanismo de barrido mueve la sonda por encima de la superficie, se ajusta de modo automático la altura de la sonda para mantener constante la corriente del efecto túnel. Estos ajustes minúsculos permiten dibujar las ondulaciones de la superficie. Después de muchas pasadas hacia adelante y hacia atrás se utiliza una computadora para crear una representación tridimensional del material.
Otro tipo de microscopio de sonda de barrido es el microscopio de fuerza atómica (Atomic Force Microscope, AFM), que no emplea la corriente de efecto túnel y que por lo tanto puede utilizarse también en materiales no conductores. A medida que la sonda se mueve a lo largo de la superficie de la muestra los electrones de la sonda de metal son repelidos por las nubes electrónicas de los átomos de la misma. La altura de la sonda se ajusta de modo automático para mantener constante la fuerza recibida. Un sensor registra el movimiento ascendente y descendente de la sonda y entrega la información a una computadora, que a su vez la utiliza para dibujar una imagen tridimensional de la superficie del espécimen.
Partes más importantes de un microscopio óptico
Ocular (Ocular): Qué el utilizador mira a través para examinar el espécimen. La potencia del ocular multiplicada por la potencia objetiva iguala la ampliación total (es decir 10X objetivo 10X del ocular X = 100X. El espécimen se ha agrandado 100 veces).
Cuerpo: El microscopio puede venir como un monocular, (un ocular y tubo), o binocular, (dos oculares y tubos). Si es un binocular generalmente solamente un tubo del ocular será ajustable mientras que el otro es fijo.
Objetivos: El objetivo es la lente más importante del microscopio para producir una imagen clara de la alta resolución. El objetivo tiene varias funciones importantes. Debe recolectar la luz que viene de cada uno de las varias partes o puntas del espécimen. Debe tener la capacidad de reconstituir la luz que viene de las varias puntas del espécimen en las varias puntas correspondientes de la imagen. El objetivo se debe construir de modo que sea enfocado cerca bastante al espécimen para proyectar una imagen magnificada, verdadera para arriba en el tubo del cuerpo.
Etapa mecánica: Un dispositivo para llevar a cabo diapositivas con seguridad con las perillas con estrías separadas para mover la diapositiva desde frente a la parte posterior (norte y al sur) o de lado a lado (este y al oeste). Estas perillas pueden estar en los ejes separados o en un eje coaxial. Pueden enderezar o zurdo.
Condensador del sub-stage: Se cabe debajo entre de la etapa del microscopio, la lámpara que ilumina y el espécimen. La abertura del condensador y el enfocarse apropiado del condensador son de importancia crítica en realizar la capacidad máxima de la lente objetiva en uso. Asimismo, el uso apropiado del diafragma ajustable del diafragma de la abertura (incorporado en el condensador) es también importante para asegurar la iluminación apropiada y contraste. La apertura y el cierre del diafragma del diafragma controla el ángulo de los rayos de la iluminación que pasan a través del condensador, a través del espécimen y entonces en la lente objetiva.
Controles del foco de Fine/Coarse: También suele llamárseles tornillos macrométrico y micrométrico, en ambas caras del soporte del microscopio hay dos conjuntos de perillas del ajuste. La perilla del ajuste aproximado para movimientos que se enfocan incrementales más grandes y la perilla del ajuste fino para movimientos que se enfocan incrementales más pequeños. Las perillas del ajuste sirven para traer el objetivo y el espécimen más cercano juntas o para engendrar aparte. En la mayoría de los microscopios las perillas del ajuste levantan o bajan la etapa; en algunos microscopios las perillas levantan o bajan el tubo el microscopio o el pedazo de la nariz.
Iluminador (Estándar): Esta parte proporciona a la iluminación requerida para realizar cualquier función con el microscopio. Es básicamente una fuente de alimentación electrónica que proporciona a electricidad a la fuente de la lámpara. Esta fuente de la lámpara podía ser una lámpara estándar del tungsteno o lámpara del halógeno. **time-out** ventaja halógeno tipo lámpara ser más luz para publicar salida vatiaje y en bajo nivel iluminación luz no tender para dar vuelta amarilla a medida que estándar tungsteno lámpara hacer.
Cubierta Del Iluminador (Estándar): Aquí es adonde el elemento electrónico asociado al iluminador se pone. Algunos microscopios más viejos tienen transformadores separados del control y el único interior de la cubierta es la lámpara, ensamblaje del socket, condensador del campo y el diafragma del diafragma del campo.
Diafragma del campo Condenser/Iris: Esta pieza del microscopio contiene la lente de condensador de la lámpara y el diafragma del diafragma del campo. Este diafragma controla el área del círculo de la luz que ilumina el espécimen.
Tejidos vasculares o conductores. Haces
Una porción del sistema vascular primario del tallo o de la hoja, constituye un cordón o haz vascular.
El floema y la xilema se hallan asociados no sólo en su conjunto, sino también por lo regular dentro de sus partes, es decir, en los haces vasculares. Las diferentes maneras de disponerse los tejidos vasculares dentro de los haces ha permitido clasificarlos en distintos tipos:
Haz colateral abierto:
Es aquel que conserva algunas células meristemáticas (o procambiales) entre el floema (líber) y la xilema (leño), productoras de elementos secundarios con origen primario.
Estas células procambiales constituyen una zona generatriz, integrada por células meristemáticas situadas entre el floema y la xilema, la cual por repetidas divisiones tangenciales de sus células produce xilema secundaria hacia la parte interna y floema secundario hacia el exterior. Poseen cambium en general las Gimnospermas y las Dicotiledóneas y su actividad se debe al crecimiento secundario de los mismos. El cambium que se halla entre la xilema y el floema se llama cambium fascicular.
Haz colateral cerrado:
Se considera cerrado el haz donde el floema y la xilema están en contacto inmediato, incapaces de modificar su estructura primitiva. Este tipo de haz es característico de las plantas monocotiledóneas. En estos tipos de haces todas las células del procámbium se diferencian en elementos de la xilema y el floema y ninguno permanece en estado meristemático como aquellos del cambium fascicular de las Gimnospermas y Dicotiledóneas. Debido a la ausencia del cambium, la mayoría de las monocotiledóneas no experimentan crecimiento en grosor.
En los haces conductores existen generalmente vainas de tejidos mecánicos en el lado exterior del floema y, a veces también en lado interno del haz, por fuera de la xilema; estas vainas en sección transversal aparecen como extractos semilunares de esclerénquima. Son pues, incompletos en los haces abiertos. A cada lado del haz, es decir, junto al límite entre la xilema y el floema, la vaina se convierte en una banda de células parenquimáticas más débilmente engrosadas, que permiten el intercambio de agua y materias alimenticias entre el haz y el parénquima circundante.
Haz bicolateral:
Se presentan en determinadas familias de plantas como Cucurbitáceas y Solanáceas; estas poseen dos zonas de floema: una en posición normal, como en el haz colateral y la otra más hacia adentro que el leño y menos desarrollado que la primera.
Haz concéntrico:
En algunas Pteridofitas y Monocotiledóneas el tipo de haz es denominado concéntrico; los hay de dos clases atendiendo a la posición de los tejidos vasculares. Con la xilema en la parte externa rodeando el floema, (tipo frecuente en los rizomas de las Monocotiledóneas), llamándose por ello concéntrico perixilemático. Y con la xilema en la parte central y el floema al exterior (frecuentemente se presenta en los helechos, por ejemplo, en el género Pteris), se designa con el nombre de concéntrico perifloemático.
Haz radial:
Se encuentra en la raíz y es característico de este órgano. Este haz conductor radial comprende varios cordones xilemáticos y floemáticos, que se disponen como los radios de una rueda, alternando los de una clase con lo de la otra. El conjunto de los cordones xilemáticos de ordinario aplanado lateralmente, presenta pues forma de estrellados en la sección transversal. Los entrantes comprendidos entre las láminas leñosas están ocupados por los Cordones cribosos, que quedan separados de los cordones xilemáticos por una o varias capas de células parenquimáticas.
En el haz radial, la formación de todos los elementos de los cordones xilemáticos y floemáticos avanzan progresivamente de la periferia del haz hacia el centro del mismo, en la porción más externa quedan por ello las células primiciales o primarias (el protoxilema y protofloema); les sigue hacia el interior los elementos diferenciados más tarde (metaxilema y metafloema). Estos haces son típicos de raíces; en el vástago se presenta raramente, por ejemplo, en algunos helechos. El número de radios xilemáticos varía de unas especies a otras.
Parénquima reservante.
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas, presenta funciones metabólicas grandes vacuolas.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
De acuerdo con su diferenciación funcional se puede distinguir: parénquima de relleno, clorofílicos, reservantes, acuífero y aerífero.
Tejidos parenquimáticos. Relleno
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
La función principal del parénquima depende del tipo de especialización que presente, y podría mencionarse el almacenamiento y producción de sustancias alimenticias, transporte de sustancias célula a célula, producir raíces adventicias, reparar daños de la corteza, excreción y secreción, etc.
También, en muchos órganos (ej. hojas con estructura isolateral, tallos y frutos), tejidos especializados, como los vasculares, (floema), aparece en forma de tejidos parenquimáticos pocos diferenciados, en los que actúa como un tejido de relleno, brindándole solidez al órgano o tejido en cuestión.
En el cuerpo de la planta el parénquima fundamental constituye la masa en la que se encuentran incluidos todos los demás tejidos. Gracias a la turgencia de sus células sirve para dar solidez general al cuerpo vegetativo, de hecho, etimológicamente, la palabra parénquima significa tejido que está en medio (tejido de relleno).
Este tejido se carácteriza por:
Tallos
Es la porción del eje de la planta que sostiene las hojas, se desarrolla a partir de la plúmula del embrión y se caracteriza por la presencia de nudos y entrenudos. En cada nudo se encuentra una o varias hojas, hojas modificadas o cicatrices de hojas. La región entre dos nudos constituye un entrenudo.
En el ápice y en las axilas (de una hoja o superior a la cicatriz de ésta) se encuentran las yemas. Algunas yemas axilares pueden ser modificadas, las yemas son estructuras no desarrollada de los tallos. Se clasifican por su posición u origen (terminales, axilares, accesorias o adventicias); por su actividad (activas o latentes) y por su contenido (foliar, floral o mixto).
La función del tallo está relacionada con el transporte de agua y nutrientes a través de la planta, además sirve como sitio de reserva de asimilados y sostén de las partes aéreas.
Clases de tallos:
Desde el tiempo de los antiguos botánicos griegos como Teofrasto y hasta el siglo XVII, la división clásica de las plantas establecía, según la estructura del tallo, tres grupos: hierbas, arbustos y árboles.
Los tallos pueden ser epigeos, si se desarrollan en el aire, hipogeos o subterráneos, cuando lo hacen enterrados; en este caso se les da los nombres de rizomas, tubérculos o bulbos.
Anatomía.
Los meristemos apicales son los generadores de los primeros tejidos del tallo, de la raíz y de los apéndices (hojas). Los tejidos primarios son estructural y funcionalmente íntegros, al menos temporalmente, y conforman el cuerpo primario de la planta.
Los tejidos secundarios constituyen, el cuerpo y pueden ser agregados más tarde; son tejidos suplementarios, reemplazan o refuerzan estructural y funcionalmente ciertos tipos de tejidos primarios como la epidermis o para la protección, y los tejidos vasculares primarios, xilema y floema para la conducción.
El crecimiento primario constituye el cuerpo de la mayoría de monocotiledóneas y pteridofitas. El crecimiento secundario se agrega al crecimiento primario en gimnospermas y muchas dicotiledóneas. El crecimiento secundario se origina en el cambium felógeno que incrementan el grosor del eje.
Los tejidos permanentes del tallo son:
Las células parenquimáticas fotosintéticas contienen cloroplastos, algunas tienen leucoplastos y otras sirven de reserva de diferentes materiales como azucares, grasas y almidones.
Colénquima y esclerénquima: El esclerénquima está constituido por células con paredes secundarias, engrosadas, lignificadas o no. La función básica es mecánica. En el esclerénquima existen dos tipos reconocidos comúnmente de células, las fibras y las esclereidas. Las células de esclerénquima a diferencia de las de la colénquima contienen un bajo porcentaje de agua y en la madurez no tienen protoplasma floema y xilema:
1.- Sistema vascular primario: Los tejidos vasculares primarios del tallo corresponden al xilema y floema primarios; estos constituyen el esqueleto básico.
Difieren entre las espermatofitas en que en las gimnospermas y en la mayoría de las dicotiledóneas el cuerpo vascular primario consiste de un cilindro continuo que envuelve la medula, esto es la porción central del tallo. En las dicotiledóneas herbáceas está constituido por un número de haces estrechos y separados que forman un anillo.
La xilema y el floema de los tallos de monocotiledóneas nunca están dispuestos en capas continuas, sino en haces vasculares individuales, esparcidos a través del tallo en forma desorganizada, más que en círculo como ocurre en las dicotiledóneas que tiene sus tejidos vasculares en haces separados. El procámbium es el tejido que origina la xilema y el floema primarios. El procámbium más temprano aparece como bandas aisladas muy estrechas en el ápice del tallo; se continua hacia las zonas más desarrolladas y diferencia el tejido vascular primario que se desarrolla en forma acropetal. Las primeras células de un haz vascular que maduran corresponden al floema, luego le siguen las primeras de la xilema.
2.- Sistema vascular secundario: La disposición del tejido vascular secundario no tiene relación con el existente tejido vascular primario, la cual puede ser en forma de cilindro completo. Sin embargo, la cantidad y disposición de los tejidos secundarios, especialmente la de la xilema, puede variar desde un cilindro completo del mismo grosor, como en árboles, hasta bandas separadas como en los tallos herbáceos de algunas dicotiledóneas, caso de Cucúrbita sp.
La xilema es un tejido constituido de varios tipos de células: células no vivas relacionadas con el transporte de agua y solutos, las fibras que ayudan al fortalecimiento del cuerpo de la planta y células del parénquima para la reserva de agua.
Dentro de los elementos traqueales se diferencian dos tipos de células: traqueidas y tráqueas. Las traqueidas son células no perforadas; las áreas de contacto presentan solamente punteaduras dobles. Las tráqueas o vasos, muestran perforaciones en sus extremos, sin embargo, existen también perforaciones en las paredes laterales.
Al completar los engrosamientos y la lignificación de la pared, el protoplasto desaparece en las células de la xilema. El floema es un tejido compuesto de elementos cribosos, y células parenquimáticas especializadas llamadas anexas. Los elementos cribosos se caracterizan por presentar cribas en sus paredes, protoplasto sin núcleos y pared primaria constituida principalmente por celulosa. La función del floema es el transporte del producto de la fotosíntesis.
Tejidos parenquimáticos. Reservante
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancia de reserva. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
La función principal del parénquima consiste en almacenar sustancias, transportar sustancias célula a célula, producir raíces adventicias, repara los daños de la corteza y se encarga de la excreción y secreción.
El parénquima reservante, se encuentra en los órganos reservantes como en los cotiledones de las semillas, la médula, en la corteza de los tallos y raíces, los radios medulares, siendo, su función la de almacenar y transportar sustancias de reserva. Sus células tienen membranas sutiles, con pequeños espacios intercelulares. Almacena carbohidratos, proteínas y lípidos.
Las raíces
Órgano de las plantas superiores, casi siempre subterráneo, que desempeña varias funciones, entre ellas absorber y conducir agua y minerales disueltos, acumular nutrientes y sujetar la planta al suelo.
La raíz se diferencia del tallo por su estructura, por el modo en que se forma y por la falta de apéndices, como yemas y hojas. La primera raíz de la planta, llamada radícula, se alarga cuando germina la semilla y forma la raíz primaria.
Las raíces que se ramifican a partir de la primaria se llaman secundarias. En muchas plantas, la raíz primaria se llama pivotante, es mucho mayor que las secundarias y alcanza mayor profundidad en el suelo.
La remolacha la zanahoria son ejemplos característicos de plantas con gruesas raíces pivotantes. Algunas especies con raíces de este tipo son difíciles de trasplantar, porque la rotura de la raíz primaria determina la pérdida de casi todo el sistema radicular y la muerte de la planta.
Las raíces que brotan de los tallos se llaman adventicias. Se ven estas formaciones cerca de la base del tallo ya que se originan de cualquier parte menos de la radícala, un ejemplo de estas raíces es la que produce la yedra venenosa, estas raíces cortas tienen estructuras terminales planas que permiten fijarse.
Cuando brotan de puntos más altos, las raíces adventicias se llaman aéreas, y ayudan a sujetar la planta, como se observa en el banano, el mangle y ciertas orquidáceas.
Las plantas parásitas usan unas modificaciones de la raíz llamadas haustorios un ejemplo de esto es la cuscuta, se fija al tejido vascular de sus hospederos gracias a estas modificaciones que les permite suprimir el abastecimiento de agua y nutrientes.
Al contrario de las parásitas, estas no obtienen beneficio de la planta que lo sostiene. Además, existen plantas hemiparásitas, es decir que toman sus nutrientes del organismo huésped, y producen parte de su propio alimento.
Las raíces, de acuerdo al medio donde se desarrollan, pueden ser
Por su duración las clasificamos en
Composición
La raíz está formada por tres tipos de tejido: epidermis, o capa superficial; tejido fundamental o córtex; y estela o cilindro vascular, situado en el centro.
Algunas células de la epidermis se modifican para desempeñar la función de absorción; de ellas parten largas proyecciones tubulares llamadas pelos radicales que se sujetan a las partículas del suelo. El agua absorbida por los pelos radicales atraviesa el córtex, zona dedicada al almacenamiento de agua y nutrientes, y penetra en el cilindro vascular, que la conduce hacia el tallo.
La organización del cilindro vascular de la raíz es muy distinta de la disposición del tejido vascular del tallo. En éste, xilema y floema se agrupan en haces vasculares; por el contrario, la raíz tiene un núcleo central formado por bandas radiales de xilema que se extienden hacia el córtex externo entre las cuales se forman hileras de floema.
En las raíces aéreas, el cilindro de xilema, por lo general macizo en las raíces subterráneas, suele tener una médula central.
Desarrollo de la raíz
Las semillas de las plantas superiores contienen una planta que todavía no se desarrolla, esta es el embrión, cuando la semilla germina se observa la raíz embrionaria o radícula esta crece debido a la división de sus células y forma la Raíz primaria: Esta raíz tiene la función de posteriormente producir ramas. Raíz secundaria: Esta estructura consta de pelos absorbentes, tienen un sabor a corcho y es la parte principal de toda nuestra raíz.
Parénquima reservante. Acuífero y aerífero
El parénquima también conocido con el nombre de tejido fundamental es el que forma la masa de las plantas y efectúa las operaciones fundamentales de la vida tales como nutrición, reproducción, elaboración de sustancias de reservas. El tejido fundamental tiene las siguientes características: se encuentra en todos los órganos vegetales, es el tejido diferenciado más simple y más abundante en la planta, está formado por células redondeadas o alargadas que dejan poco espacio entre ellas, la pared primaria es delgada y está constituida por celulosa, es un tejido vivo con protoplasto muy activo que realiza la mayor parte de las plantas.
El parénquima acuífero, está constituido por células de membranas sutiles, ricas en mucílagos, en las cuales redeposita el agua por inhibición del propio mucílago. Se presentan en algunas plantas de clima seco para mantener reservas de agua en el tallo y espesor de las hojas, (plantas suculentas), como, por ejemplo, las hojas de Aloe.
El parénquima aerífero es característico de plantas palustres y acuáticas donde sus células presentan amplios espacios intercelulares, facilitando el intercambio de gases en los órganos sumergidos.
En los amplios canales aeríferos como sucede en los pecíolos de las ninfeáceas, algunas células se transforman en idioblastos, situándose a lo largo de las filas de células de dichos canales. Efectúan estos una función mecánica pues dicho tejido es excesivamente blando. Las paredes de los ideoblastos presentan cristalitos de oxalato de calcio.
Las Plantas
La conservación de la ordenación específica que caracteriza a la vida, representa un trabajo que consume energía. La vida es, pues, un proceso ligado a la posibilidad de disponer de reservas energéticas suficientes para el mantenimiento del orden estructural. En el mismo momento en que cesa la aportación de energía, se extingue la vida activa y la ordenación comienza a desmoronarse.
El organismo animal obtiene la energía necesaria para la conservación de la vida exclusivamente del alimento, las plantas verdes, en cambio, la reciben inmediatamente de la luz solar. Entre animales y plantas verdes, existe, pues una diferencia fundamental en cuanto al modo de efectuar el aprovisionamiento de energía: las plantas, si se exceptúan algunos grupos y ciertas formas muy especializadas, son autótrofas, mientras que los animales heterótrofos. Todas las demás diferencias entre la organización animal y vegetal reposan, en último término, sobre esta diferencia básica.
Las plantas verdes toman de la luz solar la energía necesaria para la asimilación del carbono y de los demás elementos, no necesitan ser móviles como los animales, que deben ir en busca del alimento: las plantas típicas, de organización elevada, permanecen fijas, enraizadas en el substrato.
Existen, sin embargo, varias excepciones de esta regla. Hay, en efecto, grupos enteros de pequeños vegetales acuáticos, en su mayoría microscópicos, que durante toda su vida nadan libremente con la ayuda de órganos locomotores especiales.
Otra consecuencia del distinto modo de nutrirse animales y vegetales es la forma, fundamentalmente distinta, según la cual se produce el crecimiento en los dos grupos: tanto los animales como las plantas necesitan grandes superficies para la realización de los procesos metabólicos, pero mientras el desarrollo de los animales conduce, por medio de la precoz invaginación de una parte de la superficie, al establecimiento de amplias superficies internas, capaces de recibir y absorber el alimento, el de las plantas se orienta precisamente al revés, en el sentido de dar la mayor amplitud posible a las superficies externas asimiladoras, que puedan ser alcanzadas por la luz. Como los vegetales no necesitan cambiar de lugar, ningún límite externo se opone a su crecimiento; a diferencia de los animales, que siempre, después de un cierto período juvenil, cesan de crecer, las plantas continúan creciendo y desarrollando nuevos órganos hasta el momento de la su muerte. Se puede decir, pues, también, que el animal tiene una forma cerrada, mientras que la del vegetal es abierta.
También existen notables diferencias entre las células que forman a los vegetales y las que constituyen a los animales.
Las células vegetales
En el mundo viviente se encuentran básicamente dos tipos de células: las procarióticas y las eucarióticas. Las células procarióticas (del griego pro, antes de; karyon, núcleo) carecen de un núcleo bien definido. Todas las otras células del mundo animal y vegetal, contienen un núcleo rodeado por una doble membrana y se conocen como eucarióticas (del griego eu, verdadero y karyon, núcleo). En las células eucarióticas, el material genético DNA, está incluido en un núcleo distinto, rodeado por una membrana nuclear. Estas células presentan también varios organelos limitados por membranas que dividen el citoplasma celular en varios compartimientos, como son los cloroplastos, las mitocondrias, el retículo endoplasmático, el aparato de Golgi, vacuolas, etc.
Los organismos procariotes son unicelulares y pertenecen al grupo de las Moneras, que incluyen las bacterias y cianobacterias (algas verde-azules).
El DNA de las células procarióticas está confinado a una o más regiones nucleares, que se denominan nucleoides, que se encuentran rodeados por citoplasma, pero carecen de membrana.
En las bacterias, el núcleo está formado por un pedazo de DNA circular de aproximadamente 1 mm de largo, torcido en espiral, que constituye el material genético esencial. Las células procarióticas son las más primitivas de la tierra, hicieron su aparición en los océanos hace aproximadamente 3,5 millones de años; mientras que las células eucarióticas fósiles tienen menos de un millón de años.
Las células procarióticas son relativamente pequeñas, nunca tienen más de algunas micras de largo y no más de una micra de grosor. Las algas verde-azules son generalmente más grandes que las células bacterianas. Así mismo, todas las algas verde-azules realizan la fotosíntesis con la clorofila a, que no se encuentra en las bacterias, y mediante vías metabólicas comunes a las plantas y algas, pero no a las bacterias.
Un gran número de células procarióticas, están rodeadas por paredes celulares, que carecen de celulosa, lo que las hace diferentes de las paredes celulares de las plantas superiores.
En la parte interna de la pared celular, se encuentra la membrana plasmática o plasmalema, la cual puede ser lisa o puede tener invaginaciones, llamados mesosomas, donde se llevan a cabo las reacciones de transformación de energía (fotosíntesis y respiración). En el citoplasma, se encuentran cuerpos pequeños, esféricos, los ribosomas, donde se realiza la síntesis de proteínas. Así mismo, el citoplasma de las células procarióticas más complejas puede contener también vacuolas (estructuras en forma de saco), vesículas (pequeñas vacuolas) y depósitos de reserva de azucares complejos o materiales inorgánicos. En algunas algas verde-azules las vacuolas están llenas con nitrógeno gaseoso.
Muchas bacterias son capaces de moverse rápidamente gracias a la presencia de flagelos.
Células vegetales y animales
Tanto las células de las plantas como las de los animales son eucarióticas, sin embargo, presentan algunas diferencias:
Las células vegetales presentan una pared celular celulósica, rígida que evita cambios de forma y posición.
Las células vegetales contienen plastidios, estructuras rodeadas por una membrana, que sintetizan y almacenan alimentos. Los más comunes son los cloroplastos.
Casi todas las células vegetales poseen vacuolas, que tienen la función de transportar y almacenar nutrientes, agua y productos de desecho. Las células vegetales complejas, carecen de ciertos organelos, como los centriolos y los lisosomas.
La fotosíntesis es el proceso mediante el cual las plantas transforman materia inorgánica en materia orgánica.
La biosfera es la capa delgada de la Tierra donde se desarrolla la vida, abarcando la…
La botánica es la disciplina estudia las plantas. Se ocupa del reino vegetal en un sentido amplio,…
La célula es la unidad estructural, funcional y biológica más pequeña de todos los seres vivos. Es considerada…
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
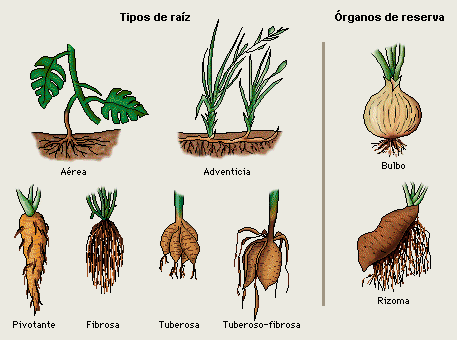{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}