Citología vegetal. La citología o biología celular es la rama de la biología que estudia las células en lo que concierne a su estructura, sus funciones y su importancia en la complejidad de los seres vivos. Citología viene del griego κύτος (célula). Con la invención del microscopio óptico fue posible observar estructuras nunca antes vistas por el hombre, las células. Esas estructuras se estudiaron más detalladamente con el empleo de técnicas de tinción, de citoquímica y con la ayuda fundamental del microscopio electrónico.
La biología celular se centra en la comprensión del funcionamiento de los sistemas celulares, de cómo estas células se regulan.
De acuerdo a la teoría celular, la célula es la unidad básica anatómica y funcional de los seres vivos. Esta teoría fue dada por Schleiden (1838) para los vegetales; por Schwann (1839) para animales y en ambos casos se determina que todos los seres vivos están constituidos por células. Existe otra teoría, la del simplasto de Hanstein (1880), la cual considera que, debido a los plasmodesmos de las células, existe continuidad del protoplasto en todo el organismo, por lo que la célula no sería la unidad básica, sino que considera como unidad al organismo en total, constituido por un simplasto (conjunto de protoplastos conectados entre sí por los plasmodesmos). Sin embargo, para los estudios anatómicos de los seres vivos. Es conveniente reconocer a la célula como la unidad básica, siguiendo la teoría celular.
Hay seres vivos formados por una sola célula y se llaman unicelulares, los pluricelulares están constituidos por numerosas células reunidas en grupos, con forma, estructura y funciones especiales, que forman los tejidos. La ciencia que se encarga del estudio de las características de las células es la citología y la que estudia los tejidos se
llama histología. Fue hasta 1667 que Robert Hooke observó por primera vez en el microscopio las células muertas de un tapón de corcho. A partir de entonces se ha ido profundizando en el estudio de la citología e histología, a la vez que se han perfeccionado diferentes tipos de microscopios como: binocular, campo claro, contraste de fases, fluorescencia, campo oscuro y electrónico; además, métodos citoquímicos de alta precisión, con lo que se ha demostrado que la célula es una estructura dinámica, capaz de aprovechar energía, sintetizar proteínas, realizar diferentes procesos metabólicos y con la presencia de ADN, efectuar su multiplicación.
Las células vegetales difieren de las de los animales en que:
a) las células de los vegetales están rodeadas de una pared rígida llamada pared celular, las de los animales no poseen pared celular;
b) la mayoría de los vegetales no pueden desplazarse, en cambio la mayoría de los animales sí se desplazan, con excepciones como anémonas y esponjas que no son fijos al sustrato y
c) los vegetales tienen la capacidad de elaborar compuestos orgánicos como azúcares, a partir de compuestos inorgánicos como agua y sales minerales, mediante el proceso de fotosíntesis y en presencia de luz y dióxido de carbono; los animales son incapaces de elaborar sus alimentos, los toman ya elaborados de las plantas o de otros animales.
El orden para hacer la descripción de las partes de la célula vegetal, es de acuerdo con su localización del exterior hacia el interior de ésta:
Pared celular
Partes de la pared celular:
Lámina media o sustancia intercelular
Pared primaria
Pared secundaria
Impregnaciones de la pared celular:
Lignina
Suberina
Sílice y carbonato de calcio
Gomas y mucílagos
Taninos y antocianinas
Puntuaciones y plasmodesmos
Protoplasto
Membrana plasmática
Inclusiones protoplasmáticas:
Hialoplasma
Núcleo
Retículo endoplasmático
Ribosomas
Dictiosomas o cuerpos de Golgi
Mitocondrias
Lisosomas
Esferosomas
Plastos (plástidos o plastidios):
Protoplastos
Cloroplastos
Cromoplastos
Leucoplastos
Inclusiones no protoplasmáticas:
Vacuolas
Sustancias ergásticas:
Grasas
Almidón
Proteínas
Taninos
Cristales
Las células vegetales exhiben gran diversidad en forma, tamaño, estructura y función. Su forma puede ser tabular (tabiques), cúbica, esférica, prismática, alargada de fibra, poliédrica, isodiamétrica, cilíndrica, estrellada, ovalada, entre otras. En cuanto al tamaño, algunas células se miden en micras (el promedio se encuentra entre 19 y 100 micras), otras se miden en milímetros y aún en centímetros, como las fibras de ciertas especies de plantas.
También las células vegetales difieren en el grosor de sus paredes y en la naturaleza del material del cual su pared celular está impregnada. En las células vivas como las que forman el tejido llamado parénquima, las paredes celulares son delgadas, ya que sus funciones son de relleno, fotosíntesis, intercambio de nutrientes y de reserva. Pero en tejidos de sostén formados por fibras, sus paredes celulares están engrosadas; también en el de conducción de agua y sales minerales llamado xilema, los vasos de este tejido están formados por elementos muertos con paredes celulares engrosadas. En las partes duras de frutos y semillas hay células muertas de sostén llamadas esclereidas, con paredes muy engrosadas.
La pared celular está compuesta en su mayoría por un carbohidrato llamado celulosa y en parte de pectosa. Las paredes engrosadas de elementos muertos pueden estar impregnadas de lignina, suberina, entre otras. Las células epidérmicas están recubiertas en su superficie externa de una sustancia grasa llama cutina.
ELEMENTOS BIOGENÉSICOS
Son elementos químicos simples que intervienen en la composición de la materia viva; son numerosos, y algunos de ellos se encuentran constantemente como:
Carbono, Hidrógeno, Oxígeno, Nitrógeno, Azufre, Fosforo, Cloro, Sodio, Potasio, Calcio, Magnesio, Hierro
Otros solo de modo eventual se hallan en algunos tejidos, órganos o formaciones especiales de animales o vegetales.
COMPUESTOS INORGÁNICOS
En este grupo se consideran el agua y las sales minerales.
El aguaes el compuesto más importan-te y el que en mayor proporción se encuentra en los organismos; la proporción varía mucho de un organismo a otro y según el medio en que vivan, pero si se puede asegurar que es el compuesto más abundante e importante sin el cual no se realizarían funciones vitales.
Las sales minerales constituyen un poco más de un 25% de las sustancias sólidas de los organismos; se encuentran disueltas en el agua y combinadas con las proteínas;
las más importantes son las sales de sodio, potasio, calcio y magnesio, en forma de cloruros, carbonatos, bicarbonatos, fosfatos, sulfatos, fluoruros, lactatos y oxalatos.
COMPUESTOS ORGÁNICOS
Los compuestos orgánicos que intervienen en la composición de la materia viva son los carbohidratos, lípidos, proteínas y ácidos nucleicos
Carbohidratos
Son compuestos que contienen hidrógeno y oxígeno, en la misma proporción que el agua, y carbono; la fórmula de la mayoría de estos compuestos se puede expresar como Cm (H2O) n
Sin embargo, estructuralmente estos compuestos no pueden considerarse como carbono hidratado, como la fórmula parece indicar.
Los hidratos de carbono son los compuestos orgánicos más abundantes en la naturaleza. Las plantas verdes y las bacterias los producen en la fotosíntesis, durante el cual absorben el dióxido de carbono del aire y por acción de la energía solar producen hidratos de carbono y otros productos químicos necesarios para que los organismos sobrevivan y crezcan.
Cuando hablamos de las características de los tejidos de las plantas tenemos que tener en mente la historia ocurrida hace unos 450 a 500 millones de años, en el paleozoico medio, cuando las plantas conquistaron la tierra. El medio terrestre ofrece ventajas respecto al medio acuático: más horas y más intensidad de luz, y mayor circulación libre de CO2. Pero a cambio las plantas tienen que solventar nuevas dificultades, casi todas relacionadas con la obtención y retención de agua, con el mantenimiento de un porte erguido en el aire y también con la dispersión de las semillas en medios aéreos. Para ello las plantas se hacen más complejas: agrupan sus células y las especializan para formar tejidos con funciones más especializadas para hacer frente a estas nuevas dificultades. Atendiendo a razones topográficas, los tejidos se agrupan en sistemas (Sachs, 1875), que se usan para resaltar la organización de estos tejidos en estructuras más amplias de la planta. Los sistemas de tejidos se agrupan para formar los órganos.
Las características de las células y tejidos de las plantas son un reflejo de su actividad y de su función:
Agua. Una de esas características es la especial relación que tienen las plantas con el agua. Todos los organismos usan agua, pero las plantas además la usan como una herramienta hidráulica. Ya que carecen de estructuras contráctiles como las células musculares, las plantas han evolucionado para aprovecharse de las propiedades físicas del agua para generar fuerzas que muevan fluidos, y generen fuerzas para el crecimiento y el movimiento. Cuando el agua se evapora por transpiración en una hoja, ese agua es reemplazado por otra agua proveniente de la hoja, y esta tira del agua del peciolo, esta de la del tallo y esta de la de la raíz, y finalmente del suelo. La diferencia de energía entre el agua de la atmósfera y la del suelo es suficiente para arrastrar agua hasta decenas de metros de altura a lo largo de los troncos. Mediante la concentración de solutos en algunos tipos celulares las plantas son capaces de generar fuerzas de expansión que producen el crecimiento celular. Se generan fuerzas tan grandes como por ejemplo para que las raíces sean capaces de romper las piedras. El movimiento de los órganos de las plantas carnívoras para atrapar a los insectos está mediada por corrientes de agua.
Autótrofas. Las plantas tienen clorofila y son fotoautotrofas. Fotoautótrofas quiere decir que utilizan la luz solar, agua y sales minerales para sintetizar todas las moléculas orgánicas que necesitan como azúcares, proteínas, lípidos y nucleótidos. Los fitocromos son capaces de percibir los cambios en el ciclo luz-oscuridad, y así anticipar floraciones o brotes. Otra característica de las plantas es que no generan prácticamente residuos resultantes de su actividad metabólica. Los animales tienen un sistema digestivo y excretor que genera constamente residuos. Por el contrario, las plantas tienen un metabolismo para generar lo que necesitan sin productos de deshecho.
Inmóviles. Como las plantas no se pueden mover, se defienden de sus predadores mediante la creación estructuras en su cuerpo como espinas, o creando sustancias tóxicas o repelentes, muchas de estas sustancias las usamos los humanos, tales como especias, colorantes, drogas, medicinas, etc. Además, tienen que competir con otras plantas por la luz, por el agua y los nutrientes, y esto determina la evolución y diversidad de sus estructuras.
Crecimiento continuo. Durante la vida de la planta hay un crecimiento continuo, lo que en las plantas con semillas ocurre en los meristemos. Al contrario que los animales, algunas plantas pueden vivir cientos de años y siempre hay un aporte de nuevas células.
Todas las células de los tejidos proceden de otras células indiferenciadas que se agrupan formando unas estructuras denominadas meristemos, aunque a veces pueden estar dispersas. Las plantas vasculares producen semillas, dentro de las cuales se forma el embrión, que se desarrolla y crece gracias a la actividad de los tejidos embrionarios o meristemáticos. A medida que la planta se desarrolla, los meristemos se mantienen en algunas partes de la planta y permiten su crecimiento, a veces a lo largo de toda la vida de la planta.
Tradicionalmente los tejidos de las plantas se agrupan en tres sistemas : sistema de protección (epidermis y peridermis), fundamental (parénquima, colénquima y esclerénquima) y vascular (xilema y floema) (Figura 1).
Figura 1. Clasificación tradicional de los tejidos de las plantas.
El sistema de protección permite resistir un medio ambiente variable y seco. Está formado por dos tejidos: la epidermis y la peridermis. Las células de estos tejidos se revisten de cutina, suberina y ceras para disminuir la pérdida de agua, y aparecen los estomas en la epidermis para controlar la transpiración y regular el intercambio gaseoso.
El sistema fundamental lleva a cabo funciones metabólicas y de sostén. Una gran proporción de los tejidos vivos de las plantas está representada por el parénquima, el cual realiza diversas funciones, desde la fotosíntesis hasta el almacén de sustancias. Para mantenerse erguidas sobre la tierra y mantener la forma y estructura de muchos órganos las plantas tienen un sistema de sostén representado por dos tejidos: colénquima y otro más especializado denominado esclerénquima. La función de mantener el cuerpo de la planta erecto pasará a los sistemas vasculares en plantas de mayor porte.
Uno de los hechos más relevantes en la evolución de las plantas terrestres es la aparición de un sistema vascular capaz de comunicar todos los órganos del cuerpo de la planta. El sistema vascular está formado por dos tejidos: xilema, que conduce mayormente agua, y floema, que conduce principalmente sustancias orgánicas en solución. Sólo hablamos de verdaderos tejidos conductores en las plantas vasculares.
Los tejidos también se pueden agrupar de otras formas. Por ejemplo, por la diversidad celular que los componen. Así, hay tejidos simples o sencillos que sólo contienen un tipo celular, como los parénquimas, mientras que otros son complejos como los de protección o conductores (Figura 2).
Figura 2. Clasificación de los tejidos de las plantas según su permanencia, capacidad de división y tipos celulares que los componen.
Los tejidos y sistemas de tejidos se agrupan para formar órganos que pueden ser vegetativos, como la raíz (órgano de captación de agua y sales), tallo (órgano para el transporte, sostén y a veces realiza la fotosíntesis) y hoja (órgano que capta la energía solar, realiza la fotosíntesis y es el principal responsable de la regulación hídrica de la planta), o bien reproductivos como la flor y sus derivados, la semilla y el fruto. Los sistemas de tejidos se distribuyen en modelos característicos dependiendo del órgano.
Antes de introducirnos en el estudio de cada uno de los tejidos y órganos tenemos que entender dos estructuras característicos de las plantas:
Pared celular
1.- Las células de las plantas presentan una estructura denominada pared celular que recubre externamente a su membrana plasmática. Se sintetiza por la propia célula y determina la forma y el tamaño de las células, la textura del tejido y la forma del órgano. Incluso los diferentes tipos celulares se identifican por la estructura de la pared. La pared celular primaria se deposita mientras la célula está creciendo o dividiéndose. La pared celular secundaria es característica de algunas células especializadas y es mayormente depositada cuando la célula ha detenido su crecimiento. Todas las células de las plantas diferenciadas contienen lamina media y pared celular primaria más o menos gruesa pero sólo unos pocos tipos celulares tienen además pared celular secundaria.
2.- A partir del estado embrionario las plantas se desarrollan y crecen gracias a la actividad de los meristemos. El primer crecimiento de todas las plantas, y único en algunos grupos, es el crecimiento en longitud. Éste se denomina crecimiento primario, y corre a cargo de la actividad de un grupo de células meristemáticas que se sitúan en los ápices de los tallos y raíces, así como en la base de los entrenudos. Estos grupos de células constituyen los meristemos primarios. Además, algunos grupos de plantas también pueden crecer en grosor, un tipo de crecimiento denominado crecimiento secundario, y lo hacen gracias a la actividad de otro tipo de meristemos denominados meristemos secundarios.
MERISTEMOS
MERISTEMOS
En el estado de embrión prácticamente todas las células de una planta pueden dividirse y diferenciarse en células funcionales. A medida que la planta se desarrolla quedan grupos de células indiferenciadas en diferentes localizaciones de su cuerpo que retienen esta capacidad proliferativa y de posterior diferenciación. En algunas ocasiones, sin embargo, se forman grupos de células con estas características por desdiferenciación a partir de células ya diferenciadas. Independientemente de cómo se formen, los meristemos son grupos de células indiferenciadas responsables del crecimiento permanente de las plantas debido a que tienen una alta capacidad de división celular y posteriormente pueden diferenciarse en una gran variedad de tipos celulares. No todas las células que se están dividiendo en un meristemo se diferencian en células funcionales, sino que algunas permanecen en estado indiferenciado tras la división mitótica, siendo éstas las que mantendrán al propio meristemo a lo largo de la vida de la planta. Los meristemos proveen de un ambiente tanto para la diferenciación como para el mantenimiento de células indiferenciadas.
Los meristemos pueden ser permanentes y ser funcionales a lo largo de la vida de la planta, como ocurre con los apicales se tallos y racíces. Otros son pasajeros y actúan durante un periodo corto de vida como los que producen las hojas, las flores, o los peciolos. También hay algunos que se pueden generar mucho después de la germinación como el felógeno y el cámbium vascular.
Las células meristemáticas presentan las características citológicas de las células indiferenciadas. Son pequeñas, isodiamétricas y tienen una pared celular primaria delgada. Su citoplasma contiene características propias, como abundantes ribosomas, un retículo endoplasmático rugoso escaso, el complejo de Golgi muy desarrollado para fabricar los componentes de la pared celular, numerosos proplastidios, una cantidad variable de pequeñas vacuolas y un citoplasma desprovisto de inclusiones. El núcleo, con mucha cromatina condensada, es grande y se sitúa en posición central. Las células meristemáticas son células totipotentes, se dividen por mitosis y posteriormente se diferencian para originar el espectro entero de tipos celulares de una planta adulta. Las plantas crecen por la producción de nuevas células, pero también por el crecimiento en tamaño de estas nuevas células debido a la incorporación de agua en sus vacuolas. En general, las células meristemáticas están densamente empaquetadas, sin dejar espacios intercelulares.
Prácticamente todos los meristemos están formados por un nicho con células madres o iniciales y éstas a su vez por células progenitoras que se dividen muchas veces y se diferencian. En las plantas el camino de diferenciación de las células meristemáticas, es decir, en qué se van a convertir, está determinado por la posición y las señales que hay en cada lugar. Aunque hay una determinación por linaje celular, la posición de la célula en el meristemo es una señal más fuerte que domina su diferenciación, es decir, en qué célula funcional se va a convertir. Las iniciales dan lugar a células que se siguen dividiendo en una zona denominada de proliferación, hasta que cesan de dividirse empiezan a elongarse, convirtiéndose entonces en zona de elnogación, para finalmente empezar la diferenciación.
La clasificación de los meristemos se realiza en base a su posición en el cuerpo de la planta y al momento en que aparecen durante el desarrollo (Figura 1).
Figura 1. Clasificación de los meristemos de las plantas.
2. Meristemos primarios
Meristemo apical caulinar Meristemo apical radical
Los meristemos primarios se originan en el embrión y son los responsables del crecimiento en longitud de la planta. Por ello a los tejidos derivados directamente de ellos también se les llama tejidos primarios. Este crecimiento en longitud se debe a que sus células se dividen sobre todo por tabicación anticlinal (ver figura). Los primeros primarios también originan órganos nuevos. Dentro de los meristemos primarios tenemos a los meristemos apicales que se sitúan en el ápice del tallo (Figura 2) y en el de la raíz principal. Estos meristemos se denominan meristemos apicales caulinares y meristemos apicales radicales, respectivamente. Del meristemo apical del tallo derivan las yemas axilares, de las cuales surgirán las ramas que llevarán en sus extremos en crecimiento meristemos apicales similares al caulinar original. Sin embargo, los meristemos apicales de las raíces laterales derivan de la endodermis de la raíz. Pero también en los ápices de las raíces laterales hay un meristemo apical caulinar. Los meristemos apicales del tallo están protegidos por los primordios foliares, mientras que los radicales lo están por la cofia o caliptra. El número de células en un meristemo apical, caulinar o radical, se mantiene relativamente constante a lo largo del tiempo puesto que hay un equilibrio entre las células producidas y las diferenciadas. En el meristemo apical caulinar, la zona central y el centro organizador forman el nicho de este meristemo. Mientras que en el meristemo radicular las células troncales forman una capa unicelular alrededor del centro quiescente. Aquí, el nicho lo forman las células del centro quiescente. A partir de cada meristemo apical se formarán la protodermis, que origina la epidermis, el procámbium que dará los tejidos conductores primarios (xilema y floema primarios), y el meristemo fundamental, que dará lugar al resto de los tejidos de la planta.
Figura 2. Meristemo apical caulinar.
La ausencia de movimiento celular en las plantas permite predecir en qué se va a convertir cada célula meristemática según la posición que ocupen en el meristemo. En los meristemos apicales, a medida que la zona central indiferenciada produce nuevas células, éstas se van alejando y entrando en la zona de influencia de otras moléculas que inician su diferenciación. La separación se va produciendo por división celular continuada.
Cámbium vascular, crecimiento secundario. Cámbium suberoso o felógeno, crecimiento secundario.
3. Meristemos secundarios
En aquellas plantas que crecen en diámetro, es decir, tienen crecimiento secundario, presentan otro tipo de meristemos denominados meristemos secundarios (Figura 1), también llamados meristemos laterales, los cuales aparecen más tardíamente en el crecimiento de la planta y son los responsables de la formación de la madera y de la corteza de los troncos. Éstos son responsables del aumento de diámetro de tallos y raíces y sus células se dividen según planos periclinales (ver figura). Estos meristemos son típicos de gimnospermas y dicotiledóneas. No aparecen en la mayoría de las pteridofitas y monocotiledóneas, ni en determinados órganos como las hojas. Por supuesto, las plantas que tienen crecimiento secundario también lo tienen primario, localizado los extremos de las raíces y ramas. Hay dos tipos de meristemos secundarios: el cambium vascular, que origina los tejidos conductores secundarios (xilema y floema secundarios), y el cambium suberoso o felógeno, que origina la peridermis. Ambos meristemos se disponen como un cilindro continuo, a veces incompleto, a lo largo del tallo o de la raíz. El cambium vascular se localiza entre el xilema secundario y el floema secundario y el felógeno entre la felodermis y el súber. El cambium vascular en los tallos procede de la unión del cambium fascicular, derivado del procámbium, más el cambium interfascicular, que se origina principalmente del parénquima. En las raíces el cambium vascular se genera a partir del procámbium que hay entre el xilema y el floema primario, así como del periciclo que rodea las crestas o costillas de xilema primario. Procámbium y periciclo se unirán para formar el cambium vascular de las raíces. A ambos lados del cambium vascular se generan unas capas de células indiferenciadas denominadas zonas cambiales. En las plantas monocotiledóneas todas las células del procámbium se convierten en xilema y floema, y no queda ninguna para convertirse en cambium vascular que produzca crecimiento secundario.
4. Otros meristemos
En las plantas monocotiledóneas la mayor parte del crecimiento en longitud del tallo no depende tanto de los meristemos apicales, sino que es responsabilidad de los meristemos intercalares. Como su nombre indica, los meristemos intercalares se encuentran intercalados entre los tejidos no meristemáticos. Los mejor conocidos son aquellos localizados en los entrenudos (principalmente próximos la base del entrenudo) y en la vaina de las hojas. Cerca del meristemo caulinar apical los nodos están muy próximos pero irán distanciándose durante el desarrollo por proliferación y elongación celular. En las dicotiledóneas el distanciamiento de los entrenudos es mayoritariamente por elongación celular.
Existen además dos meristemos adicionales relacionados con el sistema vascular: el meristemo provascular y el preprocámbium. El tejido provascular es un tejido embrionario que se sitúa en el futuro cilindro vascular. Aunque este tejido no dará directamente al xilema o al floema, sí que produce los precursores de las células que darán tanto al xilema y como al floema, es decir, dará lugar al procámbium. Las células del preprocámbium se encuentran en las hojas y son aquellas células que no se distinguen del tejido fundamental pero que son precursoras del procámbium. En las hojas las células del preprocámbium son reclutadas de una población subepidérmica de los primordios foliares. Estas células también se diferenciarán en parénquima. Estas células iniciales de la hoja se generan por la acción de la auxina a alta concentración. Se organizan en cadenas para formar los nervios de las hojas. Estos nervios no se generan por crecimiento o proliferación en longitud sino por adición de incorporación de nuevas células procambiales. Esto parece producirse por el denominado efecto de canalización de la auxina. Las células, para ser procámbium necesitan una alta concentración de auxina, y una vez diferenciadas parecen canalizar mejor la auxina a su vecina, por lo que ésta también se diferencia en procámbium, y así sucesivamente.
Pueden surgir meristemos a partir de la desdiferenciación de células como las parenquimáticas, colenquimáticas o células de la endodermis de la raíz. Por ejemplo, ya hemos visto que el cámbium vascular se forma a partir del cámbium fascicular y del cámbium interfascicular, y que el cámbium interfascicular se forma a partir de células parenquimáticas. Hay otros ejemplos de desdiferenciación. Así, todos los tipos celulares, meristemos incluidos, de las raíces laterales se forman a partir de la desdiferenciación de células de la endodermis. En los primordios de las hojas, las células del procámbium emergen del preprocámbium.
En las monocotiledóneas el crecimiento secundario se perdió durante la evolución, en todas ellas. Esto pudo ser debido a que todo el procámbium, que da lugar a los haces vasculares, se convierte en tejido y no queda meristemo. El crecimiento en grosor de los tallos se debe a crecimiento del tamaño de las células. Sin embargo, las palmeras y otras monocotiledóneas arborescentes (algunas asparagales) pueden tener tallos muy gruesos debido a un incremento en el número de células parenquimáticas y a la adición de nuevos haces vasculares en zonas alejadas del ápice del tallo. Este tipo de crecimiento se denomina a veces crecimiento anómalo y se produce por un meristemo nuevo denominado meristemos de crecimiento periférico. En las palmeras, este meristemo se encuentra externamente a los haces vasculares y se denomina meristemo de engrosamiento secundario. Este meristemo da hacia fuera células parenquimáticas y hacia dentro células parenquimáticas y células vasculares. En algunas monocotiledóneas, el meristemo periférico se extiende hacia la raíz en forma de un cilindro formado por cada vez menos células tomando aspecto de cámbium vascular. La actividad de este cilindro formará un tejido extraño similar a un tejido secundario. Este tejido sólo se produce cuando esa parte del tallo ha dejado de crecer en longitud. Curiosamente este meristemo produce la mayor parte de su tejido hacia adentro.
PARÉNQUIMA
PARÉNQUIMA
El parénquima es un tejido vivo, principal representante de los tejidos denominados fundamentales (parénquima, colénquima y esclerénquima).
Es un tejido sencillo que está implicado en una gran variedad de funciones dependiendo de dónde se encuentre, como la fotosíntesis, el almacenamiento, la elaboración de sustancias orgánicas y la regeneración de tejidos. El parénquima o las células parenquimáticas se encuentran en prácticamente todos los sistemas de tejidos de la planta. El parénquima se encuentra formando masas continuas de células en la corteza y en la médula de tallos y raíces, en el mesófilo de la hoja, en la pulpa de los frutos y en el endospermo de las semillas. Este tipo de tejido rellena espacios entre otros tejidos y dentro de ellos. Puede representar un 80 % de las células vivas de una planta. La célula parenquimática también puede aparecer asociada al xilema y floema, formando parte integral de los mismos. Parte de la capacidad de regeneración de las plantas tras sufrir heridas se debe a la actividad de las células parenquimáticas.
Está formado por un solo tipo celular, la célula parenquimática, un célula viva que generalmente presenta una pared celular primaria poco engrosada. Aunque hay ejemplos de células parenquimáticas con paredes gruesas, como las del endospermo de algunas palmeras y el caqui. Son morfológicamente muy diversas, lo que está relacionado con su función. La célula meristemática muestra menor grado de diferenciación que otras células de la planta y por eso se considera que podría ser precursora del resto de los tipos celulares durante la evolución. Es la más parecida a la célula meristemática. Normalmente hay espacios intercelulares entre las células parenquimáticas que pueden formar grandes espacios que facilitan el intercambio de gases. Las células parenquimáticas se pueden generar a de partir prácticamente todos los meristemos de la planta.
Tejido parenquimático en diferentes órganos de una planta. A. En un peciolo. B. En una hoja. C. en una raíz primaria. D. En un tallo secundario. E. En una raíz secundaria. Teñidos con azul de metileno, excepto el B que está teñido con Safranina-azul alcián.
La célula meristemática tiene la capacidad de «desdiferenciación», es decir, puede perder el grosor de su pared celular, convertirse en una célula totipotente y comenzar una actividad meristemática. Esta propiedad se usa experimentalmente para la formación de callos (masa de células indiferenciadas que es posible manipular en el laboratorio y transformar en una planta adulta). Las células parenquimáticas que están entre los haces vasculares en los tallos primarios se convierten en el meristemo cámbium vascular cuando el tallo va a empezar el crecimiento secundario.
Según su actividad y función nos encontramos 4 tipos de parénquimas:
1. Parénquima clorofílico
Este tipo de parénquima, denominado también clorénquima, está especializado en la fotosíntesis gracias a que sus células contienen numerosos cloroplastos. Se encuentra por lo general debajo de la epidermis, donde la luz llega más fácilmente, sobre todo en las hojas, aunque también es común en la zona superficial (córtex) de los tallos verdes. El clorénquima de la hoja se denomina mesófilo y se divide en dos tipos: en empalizada, más expuesto al Sol, y parénquima lagunar, en la parte más sombría. El primero tiene mayor número de cloroplastos y parece llevar a cabo una mayor tasa de fotosíntesis, estando sus células además más densamente empaquetadas. En el parénquima lagunar hay más espacios intercelulares gracias a los cuales es un buen tejido para el intercambio de gases y agua con la atmósfera.
2. Parénquima de reserva
Sus células sintetizan y almacenan diversas sustancias como azúcares en diversas formas, cristales proteicos, proteínas, lípidos, pigmentos, etc. Algunas de estas sustancias pueden encontrarse en forma sólida, aunque lo normal es que estén disueltas en la vacuola, que es el orgánulo especializado en el almacén de sustancias. También en el citoplasma y en los plastidios se pueden acumular algunas sustancias como azúcares y sustancias nitrogenadas. Algunas células almacenan un solo tipo de sustancia aunque otras pueden contener una mezcla de sustancias.
El producto de reserva más frecuente en los tejidos vegetativos (que no son semilla ni fruto) son los carbohidratos, y los almacenan de dos formas: en forma de almidón y en forma de sucrosa o sus derivados (sobre todo fructanos). El almidón se almacena en los plastidios (amiloplastos) mientras que los derivados de la sucrosa se acumulan en la vacuola. Las proteínas almacenadas suelen ser importantes como fuente de nitrógeno, un bien escaso para la célula, y generalmente el destino de estas proteínas es la degradación. Proteínas y lípidos se almacenan en el parénquima de muchas semillas, en plastidios denominados proteinoplastos y elaioplastos, respectivamente. En algunas zonas de las plantas el parénquima puede almacenar taninos y antocianinas. El color de las flores se debe a que algunas células almacenan pigmentos en los cromoplastos o en las vacuolas.
La distribución en la planta del tejido parénquimático de reserva es diversa, y puede encontrarse en la raíz, tallo, hojas y frutas. Por ejemplo, la caña de azúcar y la patata (Figura 1) almacenan material de reserva en el parénquima del tallo, y la zanahoria lo hace en el de la raíz. Otro lugar de almacenamiento es el parénquima que forma parte de los radios parenquimáticos horizontales de los haces vasculares, importantes para la reserva de los troncos en el invierno.
Figura 1. Parénquima de reserva en el tallo de la patata con amiloplastos conteniendo reservas de almidón.
3. Parénquima acuífero
Aunque todas las células parenquimáticas almacenan agua en mayor o menos medida, las células del parénquima acuífero están especializadas en esta función. Las células parenquimáticas que almacenan agua son grandes, de paredes delgadas y con una gran vacuola donde se acumula el agua. En el citoplasma o en la vacuola hay mucílagos, conjunto de sustancias que aumentan la capacidad de absorción y retención de agua. Este parénquima es característico de las plantas que viven en climas secos, denominadas plantas xerófitas. En los órganos subterráneos encargados de almacenar sustancias de reserva no suele haber tejidos especializados en el almacén de agua, aunque las células que contienen almidón u otras sustancias de reserva tienen también una gran capacidad de almacenar agua.
4. Parénquima aerífero
El parénquima aerífero o aerénquima es un tejido que contiene grandes espacios intercelulares vacíos, mayores que los normalmente encontrados en otros tejidos, por donde circulan los gases que permiten la aireación de los órganos de la planta (Figura 2).
Figura 2. Parénquima aerífero de la raíz acuática de una elodea (Elodea canadensis). Los asteriscos señalan espacios aéreos.
Este parénquima está especialmente desarrollado en las plantas que viven en ambientes muy húmedos o acuáticos (son las denominadas plantas hidrófitas), aunque también puede aparecer en algunas especies no acuáticas sometidas a estrés. Aparece tanto en raíces como en tallos. En las raíces se han descrito dos formas de producir aerénquima: esquizogenia y lisogenia. La esquizogenia es un proceso que se produce durante del desarrollo del órgano y que genera este tipo de parénquima por diferenciación celular. La lisogenia es consecuencia del estrés y las cavidades gaseosas se producen por muerte celular. El aerénquima por lisogenia se forman en plantas como el trigo, cebada, el arroz o el maíz. Algunos autores proponen un tercer tipo de formación de aerénquima denominado expansigenia en la cual se crean espacios sin que las uniones celulares desaparezcan (Figura 3 y 4).
Figura 3. Ejemplos de cómo diferentes especies crean parénquima aerífero según Seago et al. (2005). Figura 4. Distintos procesos en la formación del aerénquima (modificado de Evans, 2003).
El aerénquima es continuo desde los tallos hasta las raíces y los grandes espacios intercelulares permiten la conducción de gases, aumentando la difusión de éstos desde las hojas hasta las raíces. Esta comunicación permite a las plantas que viven en suelos húmedos o anegados mantener un nivel de oxígeno suficiente para la respiración. En suelos encharcados también es un vehículo para la liberación a la atmósfera de gases presentes en las raíces, como el etileno. Este parénquima puede considerarse como una adaptación de las plantas a la hipoxia de suelos anegados.
Las plantas con aerénquima se consideran como uno de los vehículos importantes para el paso de gases de efecto invernadero como el metano desde el suelo, pasando por la raíz y el tallo, hasta la atmósfera. Esto es particularmente importante en cultivos extensivos como los de arroz.
5. Otras células parenquimáticas
Como se mencionó anteriormente hay células parenquimáticas formando parte de otros tejidos como el xylema y el floema. En ellos hay células parenquimáticas especializadas en el transporte a grandes distancias como los elementos radiales del floema y del xilema secundarios. Además del transporte también funcionan como centros de almacenamiento. En el floema hay células parenquimáticas, denominadas acompañantes, que sirven para alimentar a las células conductoras.
Algunas células parenquimáticas están especializadas en la secreción, que incluye agua, sales, aceites, sustancias volátiles, azúcares, enzimas digestivos o sustancias defensivas. A estas células se las engloba en el denominado parénquima secretor. Algunas células parenquimáticas se especializan para formar barreras, como la endodermis, exodermis y parte de las peridermis. La exodermis aparece en las raíces de muchas angiospermas y es una capa de células en la parte más externa de la región parenquimática, justo debajo de la epidermis, y que desarrolla una pared similar a la de la endodermis. Su función es impedir la pérdida de agua. El felodermo, una capa de la peridermis, es también parénquima.
SOSTÉN
El colénquima y el esclerénquima son los tejidos especializados de sostén de las plantas. Están constituidos por células con paredes celulares gruesas que aportan una gran resistencia mecánica. A pesar de compartir la misma función, estos tejidos se diferencian por la estructura y la textura de sus paredes celulares, y por su localización en el cuerpo de la planta. En plantas de cierto porte, sin embargo, la función de soporte se lleva a cabo por los tejidos vasculares, fundamentalmente por el xilema.
1. Colénquima
Colénquima angular de una hiedra. Colénquima laminar en el tallo de un saúco.
El colénquima es un tejido vivo formado por un solo tipo celular, la célula colenquimática. Se caracteriza por estar viva, por tener paredes engrosadas y por tener una morfología elongada en la dirección del eje principal. Presentan una gruesa pared celular primaria que las diferencia de las células parenquimáticas y caracterizada por engrosamientos distribuidos de manera desigual, lo que confiere al tejido gran resistencia a la tensión y a otros tipos de estrés mecánico. Se considera pared primaria puesto que puede crecer en superficie, además de en grosor. Las células colenquimáticas, al igual que las células parenquimáticas, son capaces de reanudar una actividad meristemática gracias a que sus paredes celulares son primarias y no lignificadas, a pesar de su grosor. Es posible encontrar formas celulares difíciles de clasificar como parenquimáticas o colenquimáticas, y esto es debido a la capacidad de revertir su diferenciación que tiene el colénquima. Aunque son células vivas raramente presentan cloroplastos. Sin embargo, es un tejido transparente por lo que permite la fotosíntesis en los órganos en los que se encuentra.
Las paredes celulares de las células colenquimáticas tienen una gran cantidad de pectinas y hemicelulosas, además de celulosa. Juntos confieren a este tejido sus características de resistencia y flexibilidad. Precisamente estas características tisulares le han dado el nombre al colénquima, que deriva de la palabra griega colla, que significa goma. Si a esto le sumamos que es un tejido vivo, y por tanto con capacidad para desarrollar y engrosar sus paredes celulares, podemos decir que es el tejido de sostén por excelencia de los órganos que se están alargando, ya que tiene capacidad de adaptarse al crecimiento de cada estructura de la planta.
Colénquima lagunar del tallo de Euphorbia. Colénquima anular del tallo de una malva.
Es un tejido poco extendido en el cuerpo de las plantas ya que generalmente no está presente en las raíces, excepto en las raíces aéreas, ni tampoco en estructuras con crecimiento secundario avanzado, donde es sustituido por el esclerénquima. Está presente como tejido de soporte en órganos en crecimiento, en el tallo y hojas de numerosas herbáceas maduras, incluyendo aquellas que tienen un crecimiento secundario incipiente, y en órganos nuevos de plantas leñosas, como en tallos, hojas y partes florales de las dicotiledóneas en crecimiento. Está ausente en la mayoría de las monocotiledóneas. En los tallos y peciolos el colénquima se sitúa en posiciones periféricas, donde realiza mejor su función, bien justo debajo de la epidermis o separado de ella por una o dos capas de células parenquimáticas. Forma una especie de cilindro continuo o bien se organiza en bandas discontinuas. También existe un tipo de colénquima asociado a los haces vasculares denominado colénquima fascicular, aunque no todos los autores lo reconocen como tal.
Los distintos tipos de colénquima periférico se caracterizan por el engrosamiento de sus paredes celulares (Figura 1). Si la pared celular está engrosada de forma desigual pero no deja espacios intercelulares tenemos al colénquima angular, en el cual los engrosamientos se dan en los vértices de las células. Cuando los engrosamientos están en las paredes tangenciales externas e internas tenemos al colénquima laminar. Si el engrosamiento de la pared deja espacios intercelulares y tales engrosamientos están cerca de dichos espacios intercelulares, tenemos entonces al colénquima lagunar, o bien, si el engrosamiento es uniforme alrededor de la célula tenemos entonces al colénquima anular. Sin embargo, existen ejemplos de difícil clasificación y aparecen como formas intermedias.
Figura 1. Tipos de colénquima atendiendo a sus paredes celulares. A: Colénquima angular. El círculo rojo indica el engrosamiento en los lugares de unión de las células, sin dejar espacios intercelulares. B. Colénquima laminar. Las líneas indican el diferente grosor de las paredes celulares paralelas y perpendiculares a la epidermis. C. Colénquima anular. Las líneas indica que el grosor de las paredes celulares es más o menos homogéneo y los asteriscos los espacios intercelulares. D. Colénquima lagunar. El círculo indica que el engrosamiento de las paredes celulares ocurre en los lugares de contacto entre células, y los asteriscos que hay espacios vacíos intercelulares.
2. Esclerénquima
El esclerénquima, a diferencia del colénquima, presenta dos tipos de células con pared celular engrosada, pero ésta es secundaria y lignificada en las células maduras. La palabra esclerénquima proviene del griego «Skléros» que significa duro, seco y áspero. Las células esclerenquimáticas maduras no contienen citoplasma y son células muertas. Gracias a la estructura de sus paredes celulares el esclerénquima tiene una función muy importante en el soporte de los órganos que han dejado de alargarse. Protegen las partes más blandas de las plantas y más vulnerables a estiramientos, pesos, presiones y flexiones. Por eso, aunque está distribuido por todo el cuerpo de las plantas, ya sean estructuras con crecimiento primario o secundario, es más abundante en tallos y hojas que en raíces.
El tejido esclerenquimático es complejo. Los dos tipos de células que lo componen se distinguen principalmente por su forma, su origen y su localización. Un tipo son las fibras, células alargadas y fusiformes, y el otro las esclereidas, que son células variadas en su forma pero típicamente más isodiamétricas que las fibras. El origen de estos dos tipos celulares no está claro pero se propone que las fibras se originan por diferenciación de células meristemáticas y las esclereidas a partir de células colenquimáticas o parenquimáticas que lignifican sus paredes celulares.
Fibras de esclerénquima del maíz.
Las fibras son células alargadas de extremos puntiagudos, con una pared celular secundaria más o menos gruesa con muchas capas y con un grado de lignificación variable. Se sabe que la lignificación y la diferenciación de las fibras en los tejidos vasculares están influidos por las hormonas vegetales como las auxinas y las giberelinas, que regulan la deposición de lignina en la pared celular. La pared celular de la fibra madura puede ser tan gruesa que a veces ocupa completamente el interior celular. La mayoría de las fibras son células muertas en la madurez, aunque se han encontrado elementos fibrosos vivos en el xilema de algunas dicotiledóneas. Debido a su resistencia a la tensión son de gran importancia económica y se empaquetan por lo general formando hebras que constituyen la fibra comercial. Las fibras de las hojas de algunas monocotiledóneas son comercialmente importantes en la manufactura de la ropa y otros tejidos.
Lignina Hormonas
vegetales
Las fibras se clasifican según su posición topográfica en la planta. Las fibras extraxilares son aquellas que se encuentran en el floema (fibras floemáticas), en la corteza (fibras corticales), o bien rodeando haces vasculares (fibras perivasculares). A veces las fibras rodean el cilindro vascular en tallos con crecimiento secundario, denominándose fibras pericíclicas. Las fibras xilares se encuentran en el xilema y se clasifican en fibras-traqueidas y fibras libriformes. Ambas son alargadas con paredes gruesas, pero las fibras-traqueidas son intermediarias entre la forma de las traqueidas y las de las libriformes. Las fibras mucilaginosas o gelatinosas tienen paredes muy gruesas pero no lignificadas.
Esclereidas, de color rosado, de una hoja de un camelio.
Las esclereidas muestran paredes secundarias muy gruesas y lignificadas que a menudo están interrumpidas por unas patentes punteaduras. Sus formas pueden ser isodiamétricas, estrelladas, ramificadas, etcétera. Están ampliamente distribuidas entre las angiospermas pero son más abundantes en dicotiledóneas que en monocotiledóneas. Se encuentran en los tallos, hojas, frutos y semillas, aisladas o formando capas (Figura 2). Clásicamente se clasifican según su forma: astroesclereida, braquiesclereida, también llamada célula pétrea, macroesclereida, osteoesclereida y tricoesclereida.
Figura 2. Esclarecidas de un tallo de Ulex.
Poco se sabe de la función completa de las esclereidas. En muchos tejidos, aparte de tener una función mecánica, se les atribuye una misión protectora para paliar el efecto de los herbívoros o para disuadirlos. Aunque se han propuesto otras funciones más específicas en las hojas tales como conducir agua a la epidermis o incluso parecen ser transmisoras de luz (actúan como fibras ópticas) incrementando los niveles luminosos de las hojas. A menudo se originan tarde en la ontogenia de la planta y lo hacen por esclerosis de células parenquimáticas seguida de un crecimiento intrusivo que las hace penetrar en los espacios intercelulares del tejido donde se encuentran.
CONDUCTORES
La característica más llamativa que distingue a las plantas vasculares de las no vasculares es la presencia de tejidos especializados en la conducción de agua, sustancias inorgánicas y orgánicas. Estos tejidos son el xilema y el floema. El xilema conduce grandes cantidades de agua y algunos compuestos inorgánicos y orgánicos desde la raíz al resto de órganos de la planta, mientras que el floema conduce sustancias orgánicas producidas en los lugares de síntesis, fundamentalmente en las hojas y en las estructuras de almacenamiento, al resto de la planta.
1. Introducción
Desde el punto de vista fisiológico las plantas necesitan a los tejidos conductores para su crecimiento porque distribuyen agua y sustancias orgánicas, pero también son tejidos que hacen de soporte, a modo de esqueleto, de las partes aéreas de la planta y dan consistencia a la subterránea. Otra función de los tejidos conductores es permitir la comunicación entre diferentes partes de la planta puesto que son vías por la que viajan señales tales como las hormonas.
Durante el crecimiento primario de la planta se originan el xilema y el floema primarios. El protoxilema y el protofloema son los primeros tejidos conductores que aparecen en la planta y se forman a partir del meristemo procámbium. Esto ocurre tanto en el embrión como en las proximidades de los ápices de las plantas adultas. Posteriormente aparecen el metaxilema y el metafloema, formados también a partir del procámbium, que sustituyen paulatinamente al protoxilema y al protofloema como tejidos conductores. Si la planta tiene crecimiento secundario se forman el xilema y floema secundarios a partir del cámbium vascular, a la vez que el metaxilema y metafloema dejan de ser funcionales. El xilema y el floema, bien primarios o bien secundarios, se localizan próximos entre sí en todos los órganos de la planta puesto que proceden de la mismas células meristemáticas. El xilema y el floema están formados por distintos tipos celulares, algunos de los cuales se han usado filogenéticamente como caracteres para los estudios evolutivos.
La organización de los tejidos conductores en el tallo y la raíz es diferente. Al conjunto de haces vasculares en el tallo y en la raíz se le denomina estela, que en función de la organización reciben diferentes nombres. Por ejemplo, protoestela cuando los haces vasculares forman un cilindro sólido y sifonostela cuando los haces vasculares se disponen formando una especie de cilindro en cuyo interior hay parénquima medular (ver Figura 1).
Figura 1. Algunos de los tipos más comunes de organización de los haces vasculares, considerando la disposición del xilema y del floema. (Modificado de Furuta et al., 2014).
2. Xilema
El xilema, también llamado leño, se encarga del transporte y reparto de agua y sales minerales provenientes fundamentalmente de la raíz al resto de la planta, aunque también transporta otros nutrientes y moléculas señalizadoras. Es también el principal elemento de soporte mecánico de las plantas, sobre todo en aquellas con crecimiento secundario. La madera es básicamente xilema.
En el xilema nos encontramos cuatro tipos celulares principales. a) Los elementos de los vasos o tráqueas y b) las traqueidas constituyen los elementos conductores o traqueales. c) Las células parenquimáticas, que funcionan como células de almacenamiento o comunicación, y d) las células de sostén, que son las fibras de esclerénquima y las esclereidas.
Los elementos conductores o traqueales (tipos a y b) son células con una pared celular secundaria gruesa, dura y lignificada, y con un contenido citoplasmático que se elimina durante su diferenciación. Se distinguen a microscopía óptica por unos engrosamientos de sus paredes secundarias, los cuales pueden ser anulares, helicoidales, reticulados y punteados. El tipo de engrosamiento depende del estado de desarrollo del órgano.
Pared celular
Los elementos de los vasos (a) son células de mayor diámetro y más achatadas que las traqueidas (Figuras 2 y 3). Se unen longitudinalmente unas a otras para formar tubos llamados vasos. Por ellos el agua circula vía simplasto (por el interior de las células), y pasa de una célula a la siguiente por las perforaciones que se encuentran en sus paredes transversales (situadas en ambos extremos de la célula), denominadas placas perforadas. Estas placas pueden no aparecer el algunos vasos. Además, el agua y sustancias disueltas pueden atravesar las punteaduras areoladas situadas en las paredes laterales de la célula y pasar a otras células del xilema. Los elementos de los vasos son el principal tipo celular conductor del xilema en las angiospermas.
Figura 2. Esquema de los dos tipos de células conductoras del xilema. Las células no están a escala. Figura 3. Principales diferencias entre los elementos de los vasos y tas traqueidas
Las traqueidas (b) son el otro elemento conductor que aparece en las plantas vasculares. Las pteridofitas y gimnospermas sólo poseen este tipo traqueal como célula conductora, mientras que las angiospermas poseen tanto traqueidas como elementos de los vasos. Las traqueidas son células alargadas, estrechas y fusiformes. El agua circula por ellas y pasa de unas a otras vía simplasto atravesando las punteaduras areoladas. Estas punteaduras se encuentran sobre todo en las paredes de ambos extremos de la célula, las cuales se solapan entre células contiguas. También hay punteaduras en las paredes laterales. En general su capacidad para conducir agua es menor que la de los elementos de vasos ya que no poseen placas perforadas. Además, tienen paredes celulares más gruesas y un menor volumen interno para la conducción que los elementos de los vasos. Las traqueidas de las coníferas poseen unas punteaduras o areolas muy grandes y circulares que se caracterizan por la presencia de una estructura interna denominada toro, el cual es un engrosamiento en forma ovalada de la pared celular. El toro puede regular el flujo de agua a través de la areola.
Las células parenquimáticas (c) se organizan en los tejidos conductores de dos maneras: radialmente o axialmente. Las radiales forman filas o radios perpendiculares a la superficie del órgano, mientras que las axiales se distribuyen en grupos o tiras longitudinales en el xilema, sobre todo en el secundario (ver más adelante), y en el floema. Las células parenquimáticas radiales son células elongadas en la dirección del radio y conectadas por una gran cantidad de plasmodesmos que permiten su comunicación con otras células vecinas. En las coníferas los radios son normalmente uniseriados o biseriados, es decir, formados por una o dos filas de células, mientras que en la angiospermas son típicamente multiseriados, con muchas filas y a veces con distintos tipos de células. Los radios en el xilema se continúan con otros radios en el floema, de manera que una sola célula del cambium vascular se puede diferenciar tanto en las radiales del xilema como en las radiales del floema.
Las células parenquimáticas tienen múltiples funciones: almacén de carbohidratos como el almidón, reserva de agua, almacén de nitrógeno, hacer de comunicación entre xilema y floema, etcétera.
Las fibras de esclerénquima y esclereidas (d) tienen como función la protección y soporte (ver figura 4).
Figura 4. Esquema donde se representan los principales tipos celulares del xilema primario de una angiosperma. Xilema y floema secundarios.
El xilema primario es el primer tipo de xilema que se forma durante el desarrollo de un órgano de la planta, y está formado primero por el protoxilema y más tarde por el metaxilema. En primer lugar se forma el protoxilema a partir del meristemo procámbium. Completa su desarrollo durante la elongación del órgano y luego desaparece por fuerzas mecánicas producidas durante el crecimiento. La pared secundaria de los elementos conductores del protoxilema, las traqueidas y los elementos de los vasos, suelen tener engrosamientos anulares al inicio, para luego desarrollar otros helicoidales. El metaxilema aparece tras el protoxilema, cuando el órgano se está alargando, y madura después que se detiene la elongación. Se origina a partir del procámbium. Sus células son de mayor diámetro que las del protoxilema y las paredes celulares de los elementos conductores tienen engrosamientos en forma reticulada y posteriormente son perforadas. Es el xilema maduro en los órganos que no tienen crecimiento secundario.
Lignina
El xilema secundario se produce en aquellos órganos con crecimiento secundario a partir del meristemo cámbium vascular. Es el tejido de conducción maduro en plantas con crecimiento secundario, junto con el floema secundario.
3. Floema
El floema, llamado también líber o tejido criboso, es un tejido de conducción formado por células vivas. Su principal misión es transportar y repartir por todo el cuerpo de la planta las sustancias carbonadas producidas durante la fotosíntesis, o aquellas sustancias movilizadas desde los lugares de almacenamiento, además de otras moléculas como las hormonas vegetales.
El floema está formado por más tipos celulares que el xilema. Se compone de dos tipos de células: los elementos conductores y los no conductores. Los elementos conductores son los tubos o elementos cribosos (a) y las células cribosas (b) (Figura 5, 6, 7 y 8). Ambos tipos celulares son células vivas, aunque sin núcleo, y tienen la pared primaria engrosada con depósitos de calosa. Dentro de los elementos no conductores se encuentran las células parenquimáticas, siendo las más abundantes las denominadas células acompañantes (c). Como células no conductoras también se pueden encontrar células de soporte asociadas (d) al floema, entre las que se encuentran las fibras de esclerénquima y las esclereidas.
Figura 5. Representación esquemática de los vasos cribosos y de las células cribosas. Las células no están a escala.
Figura 6. Principales diferencias entre los vasos cribosos y las células cribosas.
Los tubos cribosos (a) (Figura 7) son típicos de las angiospermas. Son células individuales achatadas que se disponen en filas longitudinales y que se comunican entre sí mediante placas cribosas localizadas en sus paredes transversales o terminales. Las placas cribosas contienen poros de gran tamaño que comunican los citoplasmas de las células vecinas. Además, poseen áreas cribosas en las paredes laterales que son depresiones en la pared primaria con poros que atraviesan la pared completamente. Éstas sirven para comunicarse con otros tubos cribosos contiguos y con las células parenquimáticas especializadas que los acompañan llamadas células acompañantes o anexas. Los tubos cribosos constituyen el elemento conductor mayoritario en angiospermas.
Figura 7. Imagen de tubos cribosos en una angiosperma.
Las células cribosas (b) (Figura 8) son típicas de gimnospermas y pteridófitas. Son células largas y de extremos puntiagudos, comunicándose entre sí lateralmente mediante grupos de campos de poros primarios que forman las áreas cribosas. Sin embargo, no poseen placas cribosas. Se relacionan funcional y morfológicamente con una célula parenquimática especializada llamada célula albuminosa. Constituyen el único elemento conductor del floema presente en gimnospermas y pteridófitas.
Figura 8. Imagen de células cribosas en la hoja de una gimnospema.
Las células parenquimáticas (c) son células asociadas al floema. Las denominadas células acompañantes son células parenquimáticas que están estrechamente asociadas a los elementos conductores del floema puesto que mantienen metabólicamente a los tubos cribosos, ya que éstos carecen de núcleos y tienen un citoplasma reducido. Por el contrario, las células acompañantes tienen un núcleo grande y un citoplasma muy rico en orgánulos que indican una alta tasa metabólica, aunque carecen de almidón. Las células acompañantes sólo aparecen en angiospermas. En las gimnospermas las células asociadas a los elementos conductores se denominan células de Strasburguer o albuminosa, que tienen funciones similares a las acompañantes.
Otras células parenquimáticas funcionan como lugares de reserva de las sustancias transportadas por el propio floema. En algunas especies se encuentran en el floema células especializadas con función secretora. La asociación de las células parenquimáticas con los tubos o células cribosas es fuerte, cuando mueren los tubos o células cribosas también lo hacen las células parenquimáticas. En el floema primario las células parenquimáticas suelen ser alargadas y verticales, mientras que en el floema secundario tenemos un parénquima axial, con células fusiformes y alargadas, y un parénquima radiomedular con células isodimétricas.
Las fibras de esclerénquima y las esclereidas (d) se encuentran asociadas al floema con una función de protección y soporte.
Figura 9. Esquema donde se representan los principales tipos celulares del floema de una angiosperma.
El floema primario es el primer tipo de floema que aparece en los órganos en desarrollo, aparece primero como protofloema y más tarde como metafloema. El protofloema es el primer floema que aparece y se forma a partir del meristemo procámbium. El protofloema contiene elementos cribosos poco desarrollados en las angiospermas, mientras que en las gimnospermas y pteridófitas posee células cribosas, también poco desarrolladas. Las células acompañantes son muy raras o ausentes. El metafloema sustituye rápidamente al protofloema, normalmente cuando termina la elongación del órgano, y se origina también a partir del procámbium. Contiene tubos cribosos y células cribosas de grosor y longitud mayores que en el protofloema, y siempre tienen células acompañantes. Aquí aparecen las placas cribosas en los tubos cribosos. Este tejido es funcional en las plantas con crecimiento primario.
El floema secundario se forma a partir del meristemo cámbium vascular en las plantas con crecimiento secundario. En este tipo de floema los elementos conductores están muy desarrollados, así como las células acompañantes, y aparecen tanto el parénquima axial como el radiomedular. Las células del floema secundario, al contrario que en el xilema, no depositan pared celular secundaria, y son células vivas. Sin embargo, el citoplasma de los elementos cribosos puede carecer de núcleo, microtúbulos y ribosomas, y el límite entre la vacuola y el resto del citosol no es claro. En los árboles en crecimiento hay muy poco floema secundario activo implicado en la conducción de nutrientes.
PROTECCIÓN
Epidermis
Es la capa celular más externa de las plantas y se acepta que no existe en la caliptra de la raíz y que no está diferenciada en los meristemos apicales. Además, desaparece de aquellos órganos con crecimiento secundario, donde es sustituida como tejido de protección por la peridermis (ver más abajo). En las plantas que tienen crecimiento primario se mantiene a lo largo de toda la vida de la planta, excepto en algunas monocotiledóneas que la cambian por una especie de peridermo. En los tallos se origina a partir de la capa más externa del meristemo apical, o protodermis, mientras que en las raíces se origina desde las células que forman la caliptra o desde las capas más externas de la corteza parenquimática (este origen diferente hace que algunos autores denominen a la epidermis de la raíz como rizodermis). Durante el desarrollo embrionario, la superficie del embrión está formada por una capa indiferenciada denominada protodermo que se diferenciará a protodermis y ésta a la primera epidermis de la planta.
La epidermis constituye el tejido de protección de tallos, hojas, raíces, flores, frutos y semillas. Esta protección es doble, frente a patógenos y frente a daños mecánicos. Otras funciones de la epidermis trascendentales para la vida de la planta son la regulación de la transpiración, el intercambio de gases, almacenamiento, secreción, repele herbívoros, atrae insectos polinizadores, absorción de agua en las raíces, mantiene la integridad física de los órganos de la planta, protege frente a radiación solar, etcétera.
La epidermis está formada comúnmente por una sola fila de células, salvo algunas excepciones donde se aprecian disposiciones estratificadas, como es el caso de las raíces aéreas, plantas xerófitas, y en algunas hojas como las de las adelfas o los ficus. La epidermis estratificada se denomina multiseriada, múltiple, o velamen (Figura 1) cuando es en la raíz. En estas epidermis multiseriadas las capas superficiales actúan como una epidermis típica, con paredes gruesas y con una cutícula en su superficie libre, mientras que las capas profundas pueden actuar como almacén de agua.
Figura 1. Epidermis multiseriada, velamen, de la raíz aérea de una orquídea.
La epidermis está formada por diferentes tipos de células: células epidérmicas propiamente dichas o pavimentosas, células buliformes, células glandulares, células secretoras, células que componen los estomas, etcétera. Algunos tipos celulares aparecen sólo en algunas zonas de la planta como en los pétalos donde tienen pigmentos y esencias volátiles o en las de la papila del estigma donde están relacionadas con la recepción de los granos de polen.
Las células epidérmicas propiamente dichas son las más abundantes y menos especializadas, y se disponen unidas muy estrechamente, sin dejar espacios intercelulares. Tienen forma y tamaño muy variados que se suelen adaptar a la forma de la estructura que recubren. Por ejemplo, son alargadas en el tallo. En las plantas dicotiledóneas poseen paredes celulares con formas sinuosas, mientras que en monocotiledóneas suelen ser más rectas. Normalmente no tienen cloroplastos, sino plastos no clorofílicos, presentan una gran vacuola, tienen desarrollado el retículo endoplasmático y el aparato de Golgi y, por lo general, su pared celular es primaria aunque de grosor variable. Pocas veces presentan pared celular secundaria, como es el caso de las semillas, y también es poco frecuente la lignificación de las paredes secundarias, como ocurre en algunas hojas de gimnospermas. Suelen presentar en sus paredes tangenciales e internas campos de poros primarios y plasmodesmos.
Las células epidérmicas de las partes aéreas de la planta se caracterizan por sintetizar y secretar en sus superficies libres una sustancia lipídica impermeable denominada cutina, que junto con otras moléculas, se deposita en la parte externa de la pared celular para formar una capa continua llamada cutícula. La cutícula impide la pérdida de agua y la entrada de patógenos. El grosor de la cutícula varía dependiendo de la función y localización celular. A veces sobre la cutícula se depositan otras sustancias lipídicas como algunos tipos de ceras que pueden cristalizar o estar disueltas en forma de aceites. Todo esto hace que la célula sea asimétrica con una parte externa y otra interna diferentes. A veces, en la pared libre de la célula epidérmica hay unos canales, denominados ectodesmos, que permiten la comunicación del citoplasma con la cutícula y que permiten la secreción de sustancias al exterior. En la epidermis de la raíz, así como en la de los pelos radiculares, en vez de la cutina, la sustancia secretada es la suberina.
Entre las células epidérmicas propiamente dichas existen otros tipos celulares que en ocasiones pueden usarse como carácter taxonómico. Así, algunas células epidérmicas se especializan en almacenar agua, como hacen las células buliformes de las hojas de las gramíneas y otras monocotiledóneas. Estas células se caracterizan por ser mucho mayores que el resto de las células epidérmicas, por no contener cloroplastos, por poseer una gran vacuola con un alto contenido en agua y por su escasa cutícula. Parece que intervienen en el mecanismo de pliegue y despliegue de las hojas por hidratación. Se localizan en hileras paralelas a los vasos conductores o en grupos en las zonas de plegamiento de las hojas.
En la epidermis se encuentran los estomas. Las células oclusivas de los estomas son células epidérmicas especializadas que se organizan para dejar un poro u ostiolo entre ellas a través del cual se pone en contacto el medio interno de la planta con el exterior. Existe una cámara de aire bajo el ostiolo denominada cámara subestomática. Ambas estructuras, junto con las células oclusivas, forman lo que típicamente se denomina complejo estomático. Las células oclusivas tienen forma arriñonada o de palillo de tambor, presentan cloroplastos y una pared celular engrosada de manera no uniforme que posibilita que los cambios de turgencia puedan variar su morfología y de este modo aumentar o disminuir el diámetro del ostiolo.
Los tricomas o pelos también son células epidérmicas especializadas formadas a partir de células epidérmicas. Pueden ser de protección o glandulares, y a veces se usan como carácter taxonómico, es decir, sirven para clasificar especies. Los tricomas de protección pueden ser unicelulares o pluricelulares, y son especialmente abundantes en estructuras jóvenes de la planta, de las cuales pueden desaparecer cuando se hacen adultas.
Funcionalmente, los tricomas son prolongaciones epidérmicas que realizan numerosas funciones como evitar herbívoros, guiar a los polinizadores, controlar la temperatura y desecación de las hojas, o proteger frente a un exceso de luz. La mayoría de los tricomas están formados por células vivas, aunque se pueden presentar con todas sus células muertas. Las plantas que tienen muchos tricomas se denominan pubescentes.
En la epidermis de la raíz están los denominados pelos radicales, los cuales sirven para absorber agua y sales minerales. Los pelos radicales son células epidérmicas modificadas que crecen a modo de columna perpendicularmente a la superficie de la raíz. En ellos se encuentran también numerosos microorganismos simbiontes tales como las bacterias fijadoras de nitrógeno. Los pelos radicales aparecen en la zona de maduración de la raíz y se diferencian a partir de células epidérmicas indiferenciadas denominadas tricoblastos. Su patrón (número y distribución) es característico de especie, aunque también depende de las condiciones del suelo. Los tricomas tienen un origen diferente a los pelos absorbentes de la raíz, es decir, el juego de genes que se activa en cada caso es diferente.
2. Peridermis
Es un tejido de protección que sustituye a la epidermis como tejido protector en los tallos y raíces que tienen crecimiento secundario, normalmente durante el primer año con crecimiento secundario. Sin embargo, algunas plantas no suelen desarrollar la peridermis hasta varios años después de comenzar con el crecimiento secundario. La peridemis no suele aparecer en hojas ni en frutos. Su aparición aísla a la epidermis del parénquima cortical y provoca la muerte de las células epidérmicas y su descamación a medida que la raíz o tallo crecen en grosor.
La peridermis se produce por la actividad del cámbium suberoso o felógeno, un meristemo secundario y lateral que se puede originar varias veces a lo largo de la vida de la planta. Durante el primer año de crecimiento secundario se forma a partir de la desdiferenciación de las células parenquimáticas o colenquimáticas que se encuentran debajo de la epidermis, pero en algunas ocasiones también de células epidérmicas o floema primario, con lo que se puede formar un meristemo continuo o discontinuo. El primer cámbium suberoso puede durar varios años dependiendo de la especie (en el manzano, por ejemplo, más de 20 años). Más tardíamente, a veces tras varios años, el felógeno se origina en zonas más profundas a partir de células parenquimáticas del floema secundario. En las raíces el felógeno se forma a partir del periciclo. Las células de felógeno se dividen periclinalmente (ver figura) dando lugar a filas de células que se distribuyen de manera desigual hacia el interior y hacia la superficie del órgano de la planta. Las capas más externas son más numerosas y sus células se suberifican, algunas lignifican, y luego mueren formando el súber o corcho. Hacia dentro las células están vivas en una disposición apilada formando la felodermis, y aunque su forma se parece a la de las células parenquimáticas corticales, se distinguen de ellas porque se disponen en forma de hileras radiales.
Durante el crecimiento de la raíz o del tallo, en el segundo año de crecimiento secundario o posteriores se forma nuevo cámbium suberoso más internamente. Entonces, la parte externa, que puede incluir floema secundario, células parenquimáticas y la peridermis antigua, se convierte en lo que se denomina ritidoma, que es la corteza que se va desprendiendo de los árboles durante su crecimiento.
La peridermis es una capa impermeable, sobre todo el súber o corcho, al intercambio de gases entre los tejidos superficiales del tronco y raíz con el aire. Este obstáculo puede salvarse con la presencia de unas estructuras llamadas lenticelas, que permiten el intercambio de gases entre los tejidos internos del tallo y raíz y el medio ambiente. Se presentan como zonas lenticulares o circulares que sobresalen en la superficie, con un poro debajo del cual se encuentra el tejido de la lenticela, que no es más que células parenquimáticas de paredes finas y que dejan entre ellas espacios intercelulares más o menos amplios. Las lenticelas se producen con la formación de la primera peridermis y son zonas donde el cambium suberoso es más activo y deja más espacios intercelulares en los tejidos que produce. A medida que aumenta el grosor del tallo o raíz se forman nuevas lenticelas.
GLANDULARES
En las plantas es difícil discriminar entre excreción y secreción puesto que los productos de ambos procesos suelen acumularse en los mismos compartimentos vegetales o en la superficie exterior de la planta. En términos generales se puede decir que la secreción es la acumulación de metabolitos secundarios que no van a ser utilizados inmediatamente y de metabolitos primarios que van a ser utilizados de nuevo por la célula. Las células secretoras proceden de la diferenciación de otras células pertenecientes principalmente a la epidermis o al tejido parenquimático y no constituyen verdaderos tejidos. Las estructuras vegetales encargadas de la secreción tienen morfología muy diversa y localización variada. Así, se pueden encontrar en zonas internas o externas de las plantas, pueden estar constituidas por una única célula o ser pluricelulares, y además pueden producir una multitud de productos diferentes.
Los productos de desecho de las plantas son acumulados en algunas células, trasferidos a tejidos muertos o no, o a cavidades y conductos del cuerpo de la planta. Un ejemplo es la transferencia de sustancias de desecho a la madera.
Vamos a clasificar estas estructuras secretoras en función de si sus células se encuentran en la superficie de la planta o en su interior.
1. Estructuras de secreción externas
Existen multitud de estructuras secretoras en la superficie de la planta que se pueden encontrar formando pelos (tricomas), unicelulares o pluricelulares en la epidermis, o formando parte de la superficie epidérmica, y se originan a partir de células epidérmicas por división y diferenciación. Algunas secretan sustancias hidrofílicas, otras liberan sustancias lipofílicas y otras secretan pequeñas cantidades de ambas. Las secreciones son abundantes en los pétalos, donde son responsables de la fragancia de las flores, aunque también aparecen en los tallos y en las hojas. Aunque estas células están especializadas en la secreción, podría considerarse que todas las células de la epidermis son secretoras puesto que todas liberan sustancias que depositan en sus paredes o liberan al medio.
Los hidatodos son estructuras que descargan agua y algunas sustancias disueltas desde el interior de las hojas hasta su superficie. A este proceso se denomina gutación y se produce por la presión hídrica que llega desde las raíz.
Los hidatodos son modificaciones de la hoja que se localizan próximos a los bordes y a los extremos de la hoja. Estructuralmente están formados por a) traqueidas terminales, b) epitema, formado por células parenquimáticas con pocos cloroplastos y paredes delgadas, situadas al final de los terminales de los nervios, c) una vaina o envuelta que se continúa con la epidermis (las células de la envuelta pueden estar suberizadas, incluso tener bandas de Caspary), d) una abertura o poro acuoso (los poros acuáticos son pequeños estomas que han perdido la capacidad de regular su apertura y cierre). Aunque ésta es la organización general, puede haber modificaciones como la falta de envuelta, la falta de epitema, incluso puede haber aberturas sin un estoma.
Normalmente los hidatodos se encuentran en el margen y puntas de las hojas, pero en algunas especies aparecen también en la superficie y se denominan hidatodos laminares. Los hidatodos se han asociado con la liberación de agua, pero en muchas plantas serófitas se han especializado en captar agua de la condensación de la niebla, es decir, el proceso contrario. Algunos autores han descrito los denominados hidatodos activos que son aquellas estructuras liberadoras de agua pero asociadas con tricomas glandulares liberadores de agua. Éstos liberarían agua sin participación de la presión osmótica, y de ahí el nombre de hidatodos activos en contraposición con los demás que serían pasivos.
Los nectarios son estructuras secretoras productoras de azúcares resultantes de las sustancias aportadas por el floema. Se encuentran sobre todo en las flores, denominados nectarios florales, pero también en las partes vegetativas o nectarios extraflorales. Su estructura puede ser compleja o simples superficies glandulares.
Los osmóforos son estructuras secretoras que producen el olor de las plantas mediante secreción de aceites volátiles.
Las glándulas de la sal son estructuras de las plantas halófitas (viven en ambientes muy salinos) que almacenan y secretan iones para regular el contenido en sales de la planta. Las células de la sal son modificaciones de tricomas multicelulares (Figura 1). Sólo el 0.25 % de las plantas con flores se consideran halófitas. Todas las glándulas de la sal se han desarrollado para incrementar la tolerancia a la salinidad, pero como se han inventado por separado por muchas especies de plantas, sus mecanismos difieren. Son un ejemplo de evolución convergente. Las glándulas de la sal son epidérmicas y se encuentran sobre todo en las hojas de plantas que crecen en suelos salinos secos, pantanos salinos, y en los manglares. Como estas glándulas son epidérmicas en origen, se pueden considerar como tricomas especializados. La sal se mueve desde el xilema hasta el parénquima de manera apoplástica y hasta aquellas células que sustentan la glándula de la sal y que se encuentran en su base. Desde estas células la sal se mueve simplásticamente a través de plasmodesmos hasta las células almacenadoras donde se acumulan en la vacuola. Cuando se desintegra la glándula, la sal se libera formando residuos blancos en la superficie de la planta. La sal no vuelve hacia la planta desde la glándula porque las paredes de las células están cutinizadas. En tamarindos sin embargo, la sal es transferida a las células secretoras las cuales la contienen en vesículas que se fusionan con la membrana plasmática, y desde el espacio externo es transferida por pequeños poros hasta las superficie de las células.
Figura 1. Tipos de glándulas de la sal (Modificado de Dassanayake y Larkin, 2017).
Funcionalmente hay dos tipos de glándulas de la sal: aquellas que secretan la sal al exterior de la planta y aquellas que la almacenan en las vacuolas de células especializadas. Considerando su organización se clasifican en cuatro tipos: I vesícula salina (una célula grande que acumula sal); II multicelular de 4 a 40 células con células colectoras y secretoras cubiertas de cutícula; III estructuras filiformes bicelulares con una célula basal y otra apical; y IV unicelular vacuolar o filiforme secretora. Los dos primeros aparecen de dicotiledóneas y las 2 últimas en monocotiledóneas. El tipo I, vesícula salina, es una gran célula que contiene una gran vacuola que acumula sal y que posteriormente se puede romper liberando su contenido en sal. Algunas veces esta célula se encuentra encima de un grupo de células epidérmicas. El tipo II es multicelular y tienen dos poblaciones celulares, basales colectoras de sal y superficiales excretoras de sal. Las células secretoras están cubiertas de cutícula, pero, esta cutícula, o está perforada o se crea una cámara entre la cutícula y la membrana que acumula la sal excretada. Estas glándulas se organizan de forma bulbosa o en forma de disco, normalmente formando una depresión en la superficie de la hoja. La sal se suele liberar en el centro de estas estructuras. En algunas de estas glándulas suele haber un tercer tipo celular que conecta las células colectoras de las secretoras. Las glándulas del tipo III son parecidas a las glándulas secretoras de sal de las dicotiledóneas pero con estructuras más simples, con una o dos células, con o sin cutícula límite y tienen unas células basales características. El tipo IV aparece en el arroz silvestre y son pelos unicelulares.
Los tricomas glandulares son uno de los componentes de la secreción externa de las plantas (Figura 2). Por ejemplo, el olor de las flores se debe a la liberación de productos por parte de estas estructuras. Se encuentran en un 30 % de las especies. Los tricomas glandulares son normalmente multicelulares y tienen un tallo o pedúnculo, aunque algunos son unicelulares. El tallo está formado por las denominadas células sustentadoras, cuyas células más basales se anclan en la epidermis, mientras que en el extremo distal del tricoma se encuentran las células secretoras. Las células glandulares o secretoras tienen pared celular primaria, que a veces está revestida de cutícula, sobre todo en su parte de unión con las células de sostén, y un citoplasma rico en orgánulos como mitocondrias, retículo endoplasmático y sacos de aparato de Golgi. Aunque hay variaciones. Por ejemplo, las células glandulares que secretan terpenos tienen generalmente muy poco aparato de Golgi o no es visible.
Figura 2. Tricomas glandulares en una hoja de malva.
Los tricomas glandulares tienen células que secretan sustancias que liberan al medio, bien a la atmósfera o la superficie de la propia planta, con diferentes funciones. Las células glandulares de los tricomas son capaces de sintetizar sus productos de secreción, incluso algunas tienen capacidad de fotosíntesis. Es decir, actuarían como unidades independientes. Aunque en otros casos los elementos básicos para la síntesis pueden llegar a través de las células de soporte del tricoma.
Los tricomas glandulares se pueden clasificar según las sustancia que liberan o según su morfología. Hay tricomas glandulares que tienen una célula basal, una o varias de sostén y unas pocas secretoras en la punta del tricoma. Normalmente producen sustancias nada o poco volátiles que quedan en la superficie de la planta. Otros tienen una célula basal, una célula corta de tallo y una cabeza que consta de una o varias células glandulares que contienen una gran cavidad entre la cutícula y la pared celular llena con sus productos. En una misma planta nos podemos encontrar distintos tipos de estructuras secretoras, incluso dentro de un mismo órgano. Por ejemplo, en las hojas de las plantas carnívoras hay tricomas que atrapan al insecto, como los tricomas secretores de néctar o de mucílagos, y tricomas que lo digieren mediante la secreción enzimas proteolíticas. En este caso incluso un mismo tricoma puede secretar distintos tipos de sustancias. El numero de tricomas, tanto glandulares como no glandulares, ser influido por el ambiente, y se puede incrementar, por ejemplo, tras el ataque de herbívoros.
La liberación de las sustancias de secreción se puede hacer de diversas maneras. Puede haber acumulación entre la pared y la cutícula, de manera que cuando se roza el tricoma, la cutícula se desprende y las sustancias se liberan. En otras ocasiones la propia cutícula tiene poros por los que las sustancias se van liberando al exterior. Por último, algunos pelos secretores retienen las sustancias de secreción en su interior y sólo los liberan cuando la célula se rompe. Inicialmente las sustancias precursoras llegan a la célula secretora a través de las células de sostén, pero los productos de secreción, ya elaborados, no cruzan de vuelta hacia las células de sostén porque el engrosamiento posterior en el tricoma maduro de la cutícula de la célula secretora y las células barrera del propio tricoma impiden que el producto vuelva a la planta.
Hay una gran variedad de tricomas glandulares en función de las sustancias que secretan. Algunos producen y liberan ácidos orgánicos, otros secretan sal, mucílago (polisacáridos), terpenos, una mezcla de terpenos y mucílago, secretores de néctar, las plantas carnívoras tienen pelos glandulares que secretan enzimas proteolíticas, etcétera. También algunas de estas sustancias tienen efectos antimicrobianos. Una de las funciones importantes de los tricomas glandulares es mediar en la interacción entre plantas y animales. Por ejemplo, los terpenos que liberan los tricomas pueden atraer, repeler, causar alarma, o regular el desarrollo de los insectos, dependiendo de la especie. Algunos terpenos atraen a predadores o parásitos de los herbívoros. Es lo que se llama defensa indirecta.
¿De dónde consiguen los tricromas glandulares la energía y el carbono para producir sus metabolitos? Principalmente lo consiguen del cuerpo de la planta. Incluso en aquellos tricomas que tienen capacidad fotosintética, esta capacidad no se usa para fijar CO2 atmosférico sino que la energía de la fotosíntesis se emplea para otras rutas metabólicas diferentes a la de la fijación del carbono. La fuente de carbono de los tricromas glandulares son sacáridos de la propia planta, y los metabolitos propios se sintetizan en las células glandulares del tricoma.
2. Estructuras de secreción interna
Las secreciones internas son productos que se almacenarán en el interior de los tejidos de la planta, a veces durante toda la vida de ésta. Las estructuras secretoras internas se encuentran alejadas de la epidermis y se localizan principalmente en el parénquima cortical de tallos, hojas, raíces y frutos. Podemos distinguir tres tipos de estructuras secretoras:
Células secretoras. Son células aisladas que se diferencian de las células vecinas por su morfología y pueden variar desde formas isodiamétricas hasta tubos más o menos alargados. Son células muy especializadas que a menudo se denominan idioblastos. Muchas células secretoras contienen una mezcla de sustancias, y en otras el contenido no ha sido todavía identificado. Sintetizan una amplia variedad de productos que almacenan en su interior, como resinas, mucílagos, aceites, taninos e incluso sustancias cristalizables. Es frecuente ver células con precipitados critalizados en su interior que normalmente son cristales de oxalato de calcio. Esto precipitados aparecen en células aisladas en el parénquima, y también en floema secundario y en algunas células de epitelios estratificados (litocistos). Según la forma de los precipitados podemos tener drusas, en forma estrellada o esféricos (Figura 3), rafidios en forma de agregados de agujas, estiloides como cristales aciculares, y prismas. Todavía no está del todo clara la función de los cristales de oxalato de calcio en las plantas.
Figura 3. Drusas en una hoja. La imagen de arriba es un corte transversal, mientras que el de abajo es un corte en un plano horizontal y paralelo a las superficie de la hoja. Conductos resiníferos.
Las cavidades y los conductos secretores se diferencian de las células secretoras en que su producto de secreción se acumula en los espacios intercelulares. Los espacios cortos son cavidades y los largos son conductos. Estos espacios pueden localizarse en cualquier parte la planta y se forman por dos tipos de procesos: esquizógeno y lisígeno. Pueden producirse por una separación de las células que resulta en un espacio central revestido por células secretoras, denominados espacios esquizogénicos, como es el caso de los conductos resiníferos. También se pueden formar por degradación de células que previamente habían sintetizado los productos de secreción y éstos quedan en el hueco que dejan las células muertas. Dichos espacios se denominan lisogénicos, como es el caso de las cavidades lisogénicas del fruto de los cítricos. Algunos autores reconocen un tercer modelo de desarrollo, la esquizolisogénica. La formación de estas cavidades y conductos es inicialmente esquizógena (separación de células), seguida de etapas lisogénicas cuando las células que revisten el espacio sufren autolisis agrandando el espacio. Los conductos resiníferos de las escamas de las yemas de Pinus pinaster es un ejemplo de conductos que siguen el modelo esquizolisígeno. (ver figura).
Los laticíferos son células individuales o aparecen formando grupos que acumulan un líquido llamado látex. Cuando están formados por varias células se pueden organizar formando tubos. Según su estructura se pueden clasificar en articulados y no articulados, y se diferencian en que los primeros están formados por cadenas de células y los no articulados son en realidad una única célula de forma tubular y sincitial. En los no articulados, las cavidades se desarrollan desde el embrión donde hay células individuales que son las precursoras de estas estructuras. Los laticíferos articulados se originan a partir de células del meristemo apical. Sin embargo, ambos se desarrollarán en un sistema de tubos similares, cuyo grado de ramificación puede varias según la especie. Se encuentran en una gran cantidad de especies, desde herbáceas a leñosas. Las células de los laticíferos no tienen paredes lignificadas. Cuando hablamos de látex no podemos pensar en una sustancia homogénea en su composición sino que existen muchas variedades de látex. Su variación puede ser tanto en color como en composición (puede contener sustancias tales como carbohidratos, ácidos orgánicos, politerpenos, alcaloides, sales esteroles, grasas y mucílagos, y pueden tener proteínas y almidón).
TODOS los TEJIDOS
La célula
1. INTRODUCCIÓN
Esta parte del atlas está dedicada a la citología (más comúnmente denominada biología celular), y en ella vamos a estudiar la organización de la célula. Pero ¿a qué llamamos célula? La siguiente es una buena definición: una célula es la unidad anatómica y funcional de los seres vivos. Las células pueden aparecer aisladas o agrupadas formando organismos pluricelulares. En ambos casos la célula es la estructura más simple a la que consideramos viva. Hoy se reconocen tres linajes celulares presentes en la Tierra: las arqueas y las bacterias, que son procariotas unicelulares, y las células eucariotas, que pueden ser unicelulares o formar organismos pluricelulares. Las procariotas (anterior al núcleo) no poseen compartimentos internos rodeados por membranas, salvo excepciones, mientras que las eucariotas (con núcleo verdadero) contienen orgánulos membranosos internos. Uno de los compartimentos membranosos de las células eucariotas es el núcleo.
Toda célula, procariota o eucariota, es un conjunto de moléculas altamente organizado. De hecho, poseen numerosos compartimentos con funciones definidas. Vamos a considerar a un compartimento celular como un espacio, delimitado o no por membrana, donde se lleva a cabo una actividad necesaria o importante para la célula. Uno de los compartimentos presentes en todas las células es la membrana plasmática o plasmalema, que engloba a todos los demás compartimentos celulares y permite delimitar el espacio celular interno del externo.
La célula eucariota posee compartimentos internos delimitados por membranas. Entre éstos se encuentra el núcleo, delimitado por una doble unidad de membrana, en cuyo interior se encuentra el material genético, o ADN, que contiene la información necesaria para que la célula pueda llevar a cabo las tareas que permiten su supervivencia y reproducción. Entre el núcleo y la membrana plasmática se encuentra el citosol, un gel acuoso que contiene numerosas moléculas que intervienen en funciones estructurales, metabólicas, en la homeostasis, en la señalización, etcétera. Cabe destacar a los ribosomas en la producción de proteínas, al citoesqueleto para la organización interna de la célula y para su movilidad, a numerosos enzimas y cofactores para el metabolismo y a muchas otras moléculas más. Entre la membrana celular y el núcleo se encuentran también los orgánulos, que son compartimentos rodeados por membrana que llevan a cabo funciones como la digestión, respiración, fotosíntesis, metabolismo, transporte intracelular, secreción, producción de energía, almacenamiento, etcétera. Las mitocondrias, los cloroplastos, los peroxisomas, los lisosomas, el retículo endoplasmático, o las vacuolas, entre otros, son orgánulos. El citoplasma es el citosol más el conjunto de orgánulos (Figuras 1 y 2).
Figura 1. Esquema de los principales componentes de una célula animal. Figura 2. Esquema de los principales componentes de una célula vegetal.
Las células de los organismos pluricelulares están rodeados por un componente extracelular, externo a la membrana plasmática, denominado matriz extracelular. Este conjunto de moléculas está sintetizado por las propias células y es esencial para formar los tejidos, establecer las propiedades de éstos, y para modular la propia fisiología celular. En las plantas la matriz extracelular se denomina pared celular (Figura 2).
Las células procariotas, bacterias y arqueas, se definen habitualmente como células que carecen de orgánulos, al contrario que las células eucariotas. Aunque esto es cierto, en la mayoría de los casos existen procariotas que poseen orgánulos, considerando un orgánulo como un compartimento rodeado por membrana. Sin embargo, no son compartimentos aislados sino que sus membranas se continúan con la membrana plasmática, es decir, se producen por invaginación de ésta. Se han descrito al menos 4 tipos de estos orgánulos en procariotas: tilacoides, clorosomas, magnetosomas y carboxisomas.
En las siguientes páginas vamos a hacer un recorrido por el interior de la célula eucariota, pero también por sus alrededores. Algunos aspectos del funcionamiento celular no los podremos tratar con tanta profundidad como nos gustaría, como por ejemplo la expresión génica o el metabolismo celular. Ambos, por sí solos, necesitan un espacio enorme que desvirtuaría la idea que queremos dar de la célula. Existen multitud de sitios en Internet especializados en estos aspectos. Los distintos elementos que vamos a «visitar» y el orden en el que lo haremos están indicados en el menú lateral derecho.
DIVERSIDAD CELULAR
Las células son variables en forma y función. Esto fue una de las causas que hizo difícil llegar a la conclusión de que todos los organismos vivos están formados por unidades variables, pero con una organización básica común, denominadas células. La otra gran dificultad fue su tamaño diminuto.
1. Tamaño celular
El tamaño de las células se expresa en micrómetros (µm). Un micrómetro o micra es la milésima parte de un milímetro (10-3 mm), es decir, la millonésima parte de un metro (10-6 m). Una célula eucariota típica mide entre 10 y 30 µm. Esto es cierto para las células que forman parte de un gusano y para las que componen un elefante. La diferencia es que en el elefante hay más células. Para hacerse una idea de lo pequeñas que son las células imaginemos que estiramos una persona que mide 1,70 metros hasta la altura del Everest, que mide unos 8500 metros. Las células estiradas de este gigante medirían 1,3 centímetros, más pequeñas que una moneda de un céntimo de euro (sería un gigante formado por monedas de céntimo de euro).
Tamaño celular
Pero hay células eucariotas que se escapan de las dimensiones más comunes y pueden ser muy pequeñas, como los espermatozoides, cuya cabeza puede medir menos de 4 µm de diámetro, mientras que otras como los huevos de algunas aves o reptiles pueden medir más de 10 centímetros (decenas de miles de µm) en su diámetro mayor, pero sólo la yema del huevo, puesto que la clara no es parte de la célula. Piénsese en el huevo de un avestruz. Algunas células pueden tener prolongaciones de su citoplasma que miden varios metros, como sucede con las neuronas del cerebro de la jirafa que inervan las partes más caudales de su médula espinal. Más pequeñas que las células eucariotas son las células procariotas que suelen medir en torno a 1 o 2 µm de diámetro, siendo las más pequeñas los micoplasmas con dimensiones menores a 0,5 µm (Figura 1).
Figura 1. Algunos ejemplos de dimensiones celulares.
2. Número
La mayoría de los organismos vivos son unicelulares, es decir, son una única célula. Dentro de éstos son las bacterias los más abundantes, las cuales son células procariotas (anteriores al núcleo). También las especies eucariotas unicelulares son muy abundantes. Los organismos que podemos ver a simple vista son mayoritariamente pluricelulares, es decir, están formados por muchas células. Son los animales, las plantas y los hongos. En general, cuanto mayor es un organismo pluricelular más células tiene, puesto que el promedio en tamaño de las células es similar entre organismos. Hay, sin embargo, ejemplos en los que un aumento de tamaño se consigue por aumento en el tamaño celular. Las estimaciones del número de células que posee un organismo del tamaño similar al ser humano son variables y van desde 1013 (un 1 seguido de 13 ceros) hasta 1014 (un 1 seguido de 14 ceros), pero para hacerse una idea baste decir que se estima que en el cerebro humano hay unos 86.000 millones de neuronas y en el cerebro de un ratón unos 15.000 millones. Las células más abundantes del cuerpo humano son los glóbulos rojos y las neuronas del sistema nervioso. De cualquier manera, el número de células procariotas que se estima hay en la Tierra excede de largo el número de células eucariotas. Baste con decir que asociadas a nuestro cuerpo hay más células procariotas que las células eucariotas que lo componen.
3. Forma
Tipos celulares
Es común representar a las células animales con formas redondeadas pero probablemente esa sea la forma menos común que adoptan en los organismos. La morfología de las células en los tejidos animales es diversa, ¡enormemente diversa! Puede variar desde redondeada a estrellada, desde multilobulada a filiforme. También las células vegetales presentan formas variadas condicionadas por su pared celular, aunque las formas cuboidales o prismáticas son las más comunes. Véanse los siguientes ejemplos en la Figura 2. Esta variedad de formas es una de las causas por las que se tardó tanto en formular la teoría celular y en darse cuenta que todos los organismos vivos estaban formados por células con muy diversas formas y tamaños.
Figura 2. Diversas formas celulares. A) Neuronas de la corteza cerebral. B) Células musculares esqueléticas vistas longitudinalmente. C) Células vegetales de una hoja. Se puede ver la diferencia entre las células parenquimáticas, grandes y alargadas, y las de la epidermis, en la parte superior, pequeñas e irregulares. D) Distintos tipos celulares del tracto digestivo. Las células más rojizas de la parte superior son epiteliales, las alargadas pálidas de abajo son músculo liso y las verdosas situadas entre ambas son células del tejido conectivo.
4. Función
Los organismos que son una única célula son muy variados morfológicamente, dependiendo de su forma de vida y del medio al que se hayan adaptado. En estos casos, una sola célula debe realizar todas las funciones necesarias para su supervivencia y reproducción. Un organismo pluricelular, por su parte, también tiene que realizar numerosas funciones para mantener su integridad y reproducción, las cuales son llevadas a cabo por muchos tipos de células especializadas diferentes funcionando coordinadamente. Estas funciones son extremadamente complejas y variadas, desde las relacionadas con la alimentación, la detoxificación, el movimiento, la reproducción, el soporte, o la defensa frente a patógenos, hasta las relacionadas con el pensamiento, las emociones o la consciencia. Todas estas funciones las llevan a cabo células especializadas como las células del epitelio digestivo, las hepáticas, las musculares, las células germinales, las óseas, los linfocitos o las neuronas, respectivamente. La especialización supone la disponibilidad de una maquinaria molecular necesaria para su función, sobre todo formada por proteínas, que adoptan las formas más dispares para ser eficientes. Algunas funciones necesarias en un organismo pueden llevarse a cabo por células pertenecientes a un solo tipo, pero más comúnmente se necesita la cooperación de varios tipos celulares actuando de manera coordinada. Incluso, algunas funciones requieren que la célula muera tras su diferenciación como las células, que forman las uñas, o las del xylema, las cuales forman los vasos conductores de árboles plantas, que son el principal componente de la madera.
DESCUBRIMIENTO de la CÉLULA
Hoy aceptamos que los organismos están formados por células, pero llegar a esa conclusión ha sido un largo camino. El tamaño de la mayoría de las células es menor que el poder de resolución del ojo humano, que es de aproximadamente 200 micrómetros (0,2 mm). El poder de resolución se define como la menor distancia a la que se pueden discriminar dos puntos. Por tanto, para ver las células se necesitó la invención de artilugios con mayor poder de resolución que el ojo humano: los microscopios. Éstos usan la luz visible y lentes de cristal que proporcionan los aumentos. Su poder de resolución máximo es de 0,2 micrómetros, mil veces mayor que el ojo humano. Pero incluso con el uso de los microscopios se tardó en identificar a las células como unidades que forman a todos los seres vivos, debido fundamentalmente a la diversidad de formas y tamaños que presentan y también a la mala calidad de las lentes que formaban parte de los primeros microscopios.
1. Introducción
La historia del descubrimiento de la célula comienza cuando a principios del siglo XVII se fabrican las primeras lentes y el aparataje para usarlas, apareciendo los primeros microscopios. El concepto de célula está estrechamente ligado a la fabricación y perfeccionamiento de los microscopios, por tanto, a la tecnología.
Algunos de los descubrimientos y proposiciones conceptuales más relevantes en el descubrimiento de la célula se describen a continuación por orden cronológico.
2. Siglo XVII
1590-1600. A. H. Lippershey, Z. Janssen y H. Janssen (padre e hijo) son considerados como los inventores del microscopio compuesto, es decir, dos lentes de aumento colocadas cada una a un lado de un tubo. El perfeccionamiento de esta organización y de sus componentes permitiría observar más tarde a las células.
Figura 1. Dibujo hecho por R. Hooke que representa a láminas de corcho vistas al microscopio. A cada una de las estructuras huecas que forman el entramado a modo de panal de abeja les llamó celdillas o células. Apareció en Micrographia. 1664.
1610. G. Galilei describe la cutícula de los insectos. Había adaptado lentes del telescopio para inventar de manera independiente el microscopio compuesto.
1664. R. Hooke (físico, meteorólogo, biólogo, ingeniero, arquitecto) publicó un libro titulado Micrographia, donde describe la primera evidencia de la existencia de las células. Estudió el corcho y vio una disposición en forma de panal de abeja. A cada camarita la llamó celdilla o célula (Figura 1). Aunque no intuyó que aquellas celdas eran la unidad funcional de los seres vivos, la denominación de célula ha permanecido para nombrar a lo que había dentro de esas camaritas y luego se aplicó también para los descubrimientos en los animales.
1670-1680. N. Grew y M. Malpighi extendieron estas observaciones a otras plantas. N. Grew describió lo mismo que R. Hooke y a estas camaritas les llamó burbujas de fermentación (igual que en el pan). Introdujo el término de parénquima vegetal y realizó muchos dibujos de tejidos vegetales. M. Malpighi puso nombre a muchas estructuras vegetales como las tráqueas (por su similitud con las tráqueas de los insectos). Estos autores establecieron de forma detallada la organización de las estructuras microscópicas de los vegetales, que quedó bien descrita.
Las lentes eran de muy mala calidad, con grandes aberraciones cromáticas, y los microscopistas aportaban mucha imaginación. Así, G. d’Agoty consiguió ver niños completamente formados en la cabeza de un espermatozoide, el homúnculo.
A. van Leeuwenhoek
1670. A. van Leeuwenhoek construyó en la misma época microscopios simples, con una sola lente, pero con una perfección que le permitió alcanzar los 270 aumentos, más de lo que los microscopios compuestos ofrecían por aquella época. Puede ser considerado como el padre de la microbiología puesto que fue el primero en publicar observaciones de bacterias y protistas (eucariotas unicelulares). Observó gotas de agua, sangre, esperma, glóbulos rojos, etcétera. Llegó a pensar que todos los animales estaban formados por glóbulos, pero no alcanzó a asociarlos con las celdas de las plantas. Incluso, cuando se consiguieron estudiar tejidos animales con más detalle, tuvo que pasar tiempo antes de que se hiciera una asociación entre los «animalúnculos» que había descrito A. van Leeuwenhoek y las células de los tejidos animales.
En el siglo XVIII se produjeron grandes avances en el tallado de las lentes que consiguieron imágenes más nítidas. La tecnología para fabricar mejores lentes que mejoraron sustancialmente los microscopios comenzó en el siglo XVIII y continuó durante el XIX. Se atribuye a C. M. Hall (1729) el descubrimiento de un método para eliminar las aberraciones cromáticas de las lentes, es decir, defectos por descomposición de la luz al pasar por la lente. Se aplicó primero a los telescopios. De 1791 a 1806, F. Beeldsnijder y H. Van Deyl, construyeron los primeros objetivos sin aberraciones para los microscopios. En 1812, D. Brewester utiliza por primera vez objetivos de inmersión. En 1820-1837, G. B. Amici perfeccionó las lentes para microscopios, corrigiendo sus aberraciones, y diseñó objetivos con un poder de resolución y nitidez antes nunca alcanzados en los microscopios compuestos. Su diseño de los objetivos se sigue utilizando en los microscopios modernos.
3. Siglo XVIII
1759. La primera aproximación para colocar en el mismo plano a los animales y a las plantas la hizo C. F. Wolf, que dijo que existía una unidad fundamental de forma globular en todos los seres vivos. En su obra Theoria generationis argumenta con sus observaciones que los organismos vivos se forman por desarrollo progresivo y las estructuras aparecen por crecimiento y diferenciación de otras menos desarrolladas. Estas ideas eran contrapuestas a la que por aquella época existía: la teoría preformacionista, la cual proponía que los gametos llevaban organismos minúsculos ya formados y que llegaban a su estado adulto sólo por el aumento de tamaño de cada una de sus partes.
4. Siglo XIX
Figura 2. F. V. Raspail
Figura 3. Dibujo de tejido graso que aparece en Chemie organique fondé sur des méthodes nouvelles d’observation por F. V. Raspail (1833).
1820-1830. La gestación de la teoría celular comenzó en Francia con H. Milne-Edwards y F. V. Raspail (Figuras 2 y 3), que observaron una gran cantidad de tejidos de animales diferentes y publicaron que los tejidos estaban formados por unidades globulares, pero con desigual distribución. Incluyeron a los vegetales y además dieron a estas vesículas un contenido fisiológico. R. J. H. Dutrochet, también francés, escribió «si uno compara la extrema simplicidad de esta estructura chocante, la célula, con la extrema diversidad de su contenido, está claro que constituye la unidad básica de un estado organizado, en realidad, todo es finalmente derivado de la célula » (Figura 4). Estudió muchos animales y plantas y llegó a la conclusión de que las celdas de los vegetales y los glóbulos de los animales eran la misma cosa, pero con morfología diferente. Fue el primero que les asignó alguna función fisiológica. F. V. Raspail era químico y propuso que cada célula era como un laboratorio gracias al cual se organizan los tejidos y los organismos. Él dijo, y no R. Virchow, «Omnis cellula e cellula», toda célula proviene de otra célula.
Figura 4. Portada de la publicación Recherches anatomiques et physiologiques sur la structure intime des animaux et des végétaux, et sur leur motilité de R. J. H. Dutrochet (1824).
1831. R. Brown describe el núcleo. Esto es controvertido puesto que en una carta de A. van Leeuwenhoek a R. Hook en 1682 describe una estructura en el interior de los glóbulos rojos de la sangre de un pez que no podría ser otra cosa más que un núcleo, aunque no le llamó de ninguna manera. Además, en 1802, el checo F. Bauer describió una estructura celular que no podía ser otra cosa sino un núcleo.
1832. B. Dumortier describe la división binaria en células de las plantas. Detalla la aparición de la pared entre las nuevas células y propone que ese es el mecanismo de proliferación de las células.
Descubrimiento de la división celular
1835. R. Wagner describe el nucléolo.
1837. J. E. Purkinje, en Chequia, uno de los mejores histólogos de su época, propuso las ideas básicas de la teoría celular y ya dijo, no sólo que los tejidos animales estaban formados por células, sino también que los tejidos animales eran básicamente análogos a los tejidos vegetales.
1838. M. J. Schleiden, botánico alemán, formaliza el primer axioma de la teoría celular para las plantas (no estudió tejidos animales). Es decir, todas las plantas están formadas por unidades llamadas células. T. Schwann, fisiólogo alemán, hizo extensivo ese concepto a los animales y por extensión a todos los seres vivos en su publicación Mikroscopische Untersuchungen. Fue más allá diciendo que tanto células animales como vegetales estaban gobernadas por los mismos principios.
Aunque tradicionalmente se atribuye la unificación de postulados de la teoría celular a Schleiden y Schwann, hay al menos otros cuatro científicos que llegaron antes a la misma conclusión: Oken (1805), R. J. H. Dutrochet (1824), J. E. Purkinje (1834) y Valentin (1834), donde destaca R. J. H. Dutrochet (ver más arriba).
1839-1843. F. J. F. Meyen, F. Dujardin y M. Barry conectaron y unificaron diferentes ramas de la biología al mostrar que los protozoos eran células individuales nucleadas similares a aquellas que formaban parte de los animales y de las plantas, y además propusieron que los linajes celulares continuos son la base de la vida.
1839-1846. J. E. Purkinje y H. van Mohl, de manera independiente, y estudiando las células de las plantas, llaman al contenido interior de las células, excluyendo al núcleo, protoplasma. Colocar a las células vegetales y animales en el mismo plano no era frecuente en aquella época. Puesto que la idea de membrana en realidad se refería a las paredes celulares de las plantas por error, y las animales no la poseían, cuando se estudiaron con detalle células sin pared se llegó a la conclusión de que la entidad viva de la célula era el protoplasma. N. Pringsheim (1854) dijo que el protoplasma era la base material de la vida en las plantas.
1856. R. Virchow propuso a la célula como la forma más simple de manifestación viva y que a pesar de ello representa completamente la idea de vida, es la unidad orgánica, la unidad viviente indivisible. «The cell, as the simplest form of life-manifestation that nevertheless fully represents the idea of life, is the organic unity, the indivisible living One». A mediados del XIX esta teoría quedó consolidada.
La palabra «célula» y el concepto de «célula» como unidad de vida no tuvieron una buena relación durante el siglo XIX. Se había propuesto el concepto de protoplasma (Purkinje, 1839) que definía la sustancia interior de esas celdas, es decir, el citoplasma actual. Durante el siglo XIX ambas palabras compitieron para hacerse con el significado de unidad anatómica y fisiológica de los seres vivos, pero la palabra célula ganó la batalla. La palabra protoplasma ha desaparecido prácticamente de los libros de texto. Esta batalla de conceptos se produjo porque en aquella época no se tenía una idea clara de dónde residía la vida, si en el conjunto de la célula o en su interior, el protoplasma como fuerza vital.
1858. El uso de colorantes para estudiar los organismos vivos supuso un avance sin precedentes en la identificación de manera diferencial de estructuras microscópicas en los tejidos y en las propias células. Se atribuye a J. von Gerlach las primeras pruebas con soluciones de carmín en tejido nervioso. En 1829, P. Mayer introduce la tinción de hematoxilina y eosina como una tinción combinada de dos colorantes.
1879. W. Flemming describe la separación de cromosomas e introduce el término de mitosis.
Modelos de membrana
1899. C. E. Overton propone una naturaleza lipídica para la interfaz entre el protoplasma y el medio externo, y sugirió la existencia de una fina capa de lípidos rodeando al protoplasma.
5. Siglo XX
1932. Aparece el microscopio electrónico. Con él se pudieron estudiar estructuras internas de la célula que eran del orden de nanómetros (10-3 micras) (Figura 5). Un hecho que quedó resuelto con el microscopio electrónico es la existencia de la membrana plasmática rodeando a la célula, era la primera vez que se podía observar, pero también membranas formando parte de estructuras internas. El interior de la célula eucariota se mostró complejo y rico en compartimentos. Hacia 1960 ya se había explorado la célula a nivel ultraestructural.
Figura 5. Imágenes tomadas con un microscopio electrónico de transmisión. Se puede ver la capacidad de estos microscopios observando el incremento de resolución de las imágenes de izquierda a derecha. Las líneas negras de la imagen de la derecha corresponden a las membranas celulares.
TEORÍA CELULAR
La teoría celular sintetiza los principales descubrimientos citados en el apartado anterior en los siguientes postulados:
1.- La unidad estructural y funcional de los seres vivos es la célula.
2.- Todos los seres vivos están constituidos por unidades básicas denominadas células.
3.- Las células se originan exclusivamente por división de otras células.
Se puede añadir que las células pueden vivir de forma aislada, constituyendo seres unicelulares, o como parte de organismos complejos pluricelulares. En este último caso, las células se asocian formando poblaciones que se reparten las funciones del organismo, especializándose cada tipo celular en una o varias misiones determinadas.
Siendo estrictos, uno de estos postulados está formulado de manera incompleta: «toda célula procede de otra célula». Como veremos en el siguiente apartado, la teoría sobre el origen de la vida es la teoría del origen de la célula, y en ella se sostiene que las primeras células aparecieron gracias a procesos físico-químicos. Por tanto, podríamos reformular este postulado diciendo que toda célula procede de otra célula, excepto las primeras células en el origen de la vida.
Un avance que también puede hacer reformular el postulado 3 viene del campo de la biología sintética. Se han realizado experimentos en los que se ha sintetizado un genoma bacteriano completo en el laboratorio, y se ha incluido en otra bacteria a la que previamente se le había eliminado su propio ADN (Gibson et al., 2010). El resultado es una célula producida en el laboratorio, aunque sólo el ADN se ha sintetizado químicamente. Sin embargo, puede ser el primer paso hacia una síntesis artificial en el laboratorio de una célula completa exclusivamente a partir de moléculas orgánicas sencillas. Recientemente se ha sintetizado un cromosoma eucariota completo (Annaluru et al., 2014).
ORIGEN DE LA CÉLULA
El problema del origen de la vida es el problema del origen de la célula. No se sabe cómo apareció la primera célula en la Tierra, pero se acepta que su origen fue un fenómeno físico-químico. Esta visión llegó con las propuestas de A.I. Oparin y J.B.S. Haldane en torno a los años 20 del siglo pasado (también fue sugerida por C. Darwin en una carta personal).
Puesto que es un proceso físico-químico surgen dos posibilidades interesantes en el campo de la biología. a) Podemos crear vida. Se podría «fabricar» una célula utilizando las moléculas que existen hoy en día en las células actuales y colocándolas todas juntas dentro de una vesícula membranosa. b) Vida extraterrestre. Existe la posibilidad de que en otro lugar del Universo se hayan dado las condiciones necesarias, similares a las que se dieron en la Tierra, para la aparición de la vida extraterrestre.
1. ¿Qué es un ser vivo?
Para investigar el origen de la vida deberíamos saber reconocer a un ser vivo. Podemos decir que es un organismo que tiene la cualidad de la vida. Pero ¿qué es la vida? Actualmente se tiende a no proponer una definición sino a considerar a la vida como un conjunto de propiedades que debería poseer un organismo para ser considerado como vivo. Se suelen incluir:
a) Reproducción o transmisión de información codificada por el ácido desoxirribonucleico o ADN.
b) Mantenimiento de la homeostasis interna gracias a su capacidad para obtener energía externa (metabolismo).
c) Tener capacidad para producir respuestas a estímulos externos o internos.
d) Evolución condicionada por la interacción con el medio externo, capacidad para la adaptación (evolución darwiniana).
e) Etcétera.
2. ¿Dónde aparecieron las primeras células?
Aunque se acepta que la formación de las primeras células ocurrió en la Tierra a partir de moléculas orgánicas que existían en el agua, hoy en día no se descarta que parte de las moléculas orgánicas que se necesitaron para crear la vida se sintetizaran en otros planetas o en el propio espacio. Algunos meteoritos presentan una gran cantidad de materia orgánica, incluyendo algunas de relevancia biológica como aminoácidos, nucleótidos y azúcares. La teoría de la panespermia (literalmente, semillas en todas partes) postula un origen extraterrestre de la vida o de las «semillas» de la vida que llegaron a la Tierra. Por tanto, sería plausible la existencia en otros planetas de organismos similares a los de la Tierra.
3. ¿Cuándo aparecieron las primeras células?
La Tierra se formó hace unos 4.500 millones de años. Los indicios fósiles sugieren que los primeros seres orgánicos que dejaron huellas aparecieron entre 3500 y 3800 millones de años atrás (Figura 1). El proceso físico-químico de formación de estos primeros organismos debió empezar antes, en una etapa denominada prebiótica.
Figura 1. Secuencia temporal aproximada de la aparición de la vida en la Tierra y algunos de los organismos que emergieron después.
4. ¿Cómo aparecieron las primeras células?
Intuitivamente podemos imaginar una serie de pasos necesarios para la aparición de las primeras células a partir de sustancias químicas:
Formación de moléculas orgánicas
Las células están formadas por moléculas orgánicas, además del agua e iones. Las principales son proteínas, nucleótidos, azúcares y grasas. ¿Cómo se formaron? a) Condiciones físicas extremas. Si se coloca en un matraz una disolución acuosa con sustancias como CO2, amoniaco, metano e hidrógeno, y se les somete a una alta temperatura y a descargas eléctricas, se consigue que se formen pequeñas moléculas orgánicas como cianuro de hidrógeno, formaldehído, aminoácidos, azúcares, purinas y pirimidinas. Éste fue el experimento que realizaron Miller y Urey intentando simular las condiciones primitivas (Figura 2). No demuestra que estas moléculas se formaran así en el origen de la vida, pero es una prueba de que se pueden formar mediante reacciones físico-químicas. Hoy se tiende a situar esa síntesis prebiótica en los alrededores de las fumarolas, donde se darían condiciones propicias y habría una cierta protección. b) Origen extraterrestre. Es seguro que las moléculas orgánicas se formaron y se siguen formando en el espacio y se encuentran en meteoritos y cometas. Es posible que gracias a cometas y meteoritos que chocaron con la Tierra de una forma masiva aportaran suficiente materia orgánica para el comienzo de la vida.
Figura 2. Esquema del sistema ideado por Miller-Urey en el que se demuestra que se pueden sintetizar moléculas orgánicas complejas a partir de otras más simples, cuando estas últimas se someten a condiciones supuestamente similares a las de la Tierra primigenia. Años 50 del siglo XX.
Formación de polímeros
Ya tenemos moléculas orgánicas, pero las más importantes para la célula suelen aparecer en forma de polímeros complejos y no como moléculas simples: las cadenas de aminoácidos forman las proteínas y los polinucleótidos forman el ADN y el ARN. Habría varias posibilidades:
a) Calor sobre compuestos secos. Hay experimentos en los cuales la aplicación de calor sobre componentes secos lleva a la aparición de polímeros orgánicos.
b) Catálisis por superficies minerales. Los minerales pueden haber sido importantes en el origen de la vida por varias razones: concentran, seleccionan, hacen de molde y catalizan reacciones de moléculas orgánicas. Los minerales podrían haber servido además como lugares de protección frente a las adversas condiciones atmosféricas y como sustratos o moldes para la polimerización y las reacciones químicas.
c) Fumarolas. El proceso de formación de moléculas orgánicas se produce hoy en día en las fumarolas, que, bajo unas condiciones de presión y calor elevados, con la ayuda de minerales, pueden producir polímeros orgánicos.
d) Fuentes hidrotermales de agua dulce. Estos serían lugares de agua dulce en contacto con fuentes volcánicas donde sería posible la hidratación-desecación constante de reductos que podrían aumentar la concentración de moléculas orgánicas y favorecer la reacción entre ellas a altas temperaturas. Este ambiente es más favorable para formar membranas espontáneamente a partir de lípidos anfipáticos que el agua de mar.
e) Membranas lipídicas. Distintos experimentos en laboratorio muestran que las membranas lipídicas, como las que hoy tienen las células, podrían ser centros de atracción, selección y concentración de moléculas simples. Esta posibilidad es interesante puesto que resolvería el problema de cómo las membranas englobaron a unas moléculas determinadas y no a otras, y cómo se llegó a la primera protocélula.
Membrana celular
Uno de los principales eventos en el origen de las células fue el desarrollo de una envuelta que aislara un medio interno y otro externo. Esto tiene muchas ventajas: a) permite tener todos los componentes necesarios próximos para las reacciones metabólicas y se hace más eficiente el proceso de replicación; b) se evita que variantes ventajosas de moléculas orgánicas sean aprovechadas por grupos competidores, es decir, egoísmo evolutivo; c) se gana una cierta independencia respecto a las alteraciones del medio externo favoreciendo la homeostasis interna. Las membranas lipídicas se producen fácilmente de forma espontánea a partir de ácidos grasos anfipáticos, es decir, moléculas que tienen una parte cargada eléctricamente y otra que es hidrófoba.
Protocélulas
Hay dos posibilidades para la asociación entre moléculas como nucleótidos y aminoácidos y las membranas (Figura 3). a) Podemos especular que estas membranas iniciales formaron pequeñas bolsas o vesículas que englobaron poblaciones de moléculas. En otro momento, debido al crecimiento de su contenido interno, estas bolsas debieron adquirir la capacidad de estrangularse y dar dos unidades hijas con características semejantes a la parental. Se producirían reacciones moleculares internas gracias a que las membranas serían permeables a moléculas pequeñas, pero no a los polímeros creados internamente, a los cuales no les sería fácil escapar. b) Otra posibilidad es que hubo una asociación inicial de moléculas orgánicas simples con membranas de lípidos. Este sistema de polímeros (oligopéptidos y oligonucleótidos) y membranas fue ganando en complejidad y dependencia hasta que algunos polímeros atravesaron la propia membrana y quedaron en su interior. Si esto fue así, cambiaría el orden de los acontecimientos puesto que las membranas serían los verdaderos protagonistas para la formación de las primeras protocélulas.
Figura 3. Modelo de «la vida fuera de la vesícula» en el que la membrana es el elemento clave para seleccionar, concentrar y favorecer las reacciones de las moléculas (modificado de Black y Blosser, 2016).
La posibilidad de que las membranas pudieran ser en realidad el elemento que disparara la atracción de moléculas y la reacción entre ellas para formar estructuras moleculares más complejos abre posibilidades sorprendentes. Las membranas son mucho más estables en aguas dulces que en aguas saladas, por lo que surge la posibilidad de que, en estos supuestos, las primeras células aparecieran en aguas dulces, en vez de en el mar.
Autorreplicación
El evento quizá más importante para pasar de la química prebiótica a conseguir una química celular fue la autorreplicación. En el modelo del mundo ARN el replicador es la molécula de ARN (ver más abajo) y en el modelo del mundo metabólico la replicación se conseguiría después de que las primeras reacciones metabólicas empezaran a funcionar.
Con la autorreplicación se consigue la propiedad de la transmisión de la información, que es una de las propiedades de la vida. Dentro de cada vesícula membranosa se crearían réplicas moleculares más o menos exactas al original. Así, diferentes vesículas membranosas se enriquecerían en ciertas variantes moleculares y competirían más eficientemente y aprovecharían más favorablemente los materiales libres. Con este proceso de competición por los recursos se emprende otra carrera que es la de la evolución darwiniana (variabilidad más selección natural), otra gran propiedad de la vida. Algunos autores proponen que no hubo una primera molécula autorreplicante sino sistemas de reacciones químicas con capacidad para aumentar el número de sus componentes moleculares y así crecer. Es decir, se replicaría el sistema de reacciones y sus componentes.
Mundo ARN
Mundo ARN. Suponiendo que el primer autorreplicante fuera una molécula, ¿qué molécula podría autorreplicarse? Todas las miradas se vuelven al ARN. Esta idea se basa en la capacidad enzimática que poseen las moléculas de ARN (denominadas por ello ribozimas). Por ejemplo, la maduración del ARNm de las células eucariotas por parte de las ribonucleoproteínas o la síntesis de proteínas en los ribosomas por parte de los ARN ribosómicos son ejemplos de actividad catalítica llevada a cabo por el ARN. No es descabellado, aunque improbable, pensar que existieran moléculas de ARN con la capacidad de unir ribonucleótidos y hacerlo con una secuencia similar de bases a la suya propia. Podrían usar como molde la complementariedad de su propia secuencia de nucleótidos. Además, la secuencia condiciona el plegamiento tridimensional de la molécula de ARN, lo que afecta a su estabilidad y a su actividad (Figura 4). Ocurrirían fallos durante la autorreplicación que producirían moléculas de ARN con distintas secuencias y por tanto con distintas propiedades. Entre ellas comenzaría una competencia darwiniana por los recursos. Por todo ello se ha propuesto que existió un mundo dominado por el ARN en la etapa prebiótica.
Figura 4. Éste es un esquema tridimensional de un ARN de transferencia existente en las células actuales. La secuencia de ribonucleótidos hace que se establezcan uniones por complementariedad de bases (trazos verdes). Esto le provoca una disposición tridimensional.
Sin embargo, en un «mundo metabólico» basado en sistemas de reacciones químicas, la replicación no sería la característica de una molécula concreta sino de todo un sistema de moléculas. Para ello se necesitaría un aislamiento del medio externo (secuestro en una vesícula membranosa), capacidad de tomar energía y moléculas del medio, crecer, dividirse y la capacidad para aumentar su complejidad de reacciones químicas.
Interacciones moleculares
Independientemente de la molécula o moléculas con capacidad de autorreplicación y competición, tendría que darse en algún momento la interacción entre moléculas diferentes (proteínas, ADN, ARN, lípidos y azúcares) y la formación de complejos y reacciones heterogéneas. Con estas interacciones se seleccionarían no ya unas pocas moléculas sino grupos heterogéneos de moléculas que actuarían en cooperación, coevolución.
Código genético
En algún momento el ARN tuvo que intervenir en la síntesis de las proteínas. Para ello hubo que inventar un código que identificara una secuencia de nucleótidos con un aminoácido determinado. Esto es lo que actualmente se denomina el código genético, en el que tres bases nucleotídicas codifican para un aminoácido determinado. Este código parece arbitrario y es prácticamente universal para todos los organismos vivientes. A estas protocélulas de las cuales partieron todas las demás células que conocemos hoy en día se les denomina LUCA (en inglés: Last universal common ancestor).
ADN
Actualmente la información que transmiten los organismos a su descendencia está codificada en forma de ADN. El ADN tiene una serie de ventajas sobre el ARN: al ser el ADN una doble hélice es más estable, es más fácil de replicar y permite reparaciones más eficientes. En algún momento de la evolución, antes de LUCA, debió darse el paso de la información desde el ARN al ADN, y quedar este último como base para la conservación, lectura y transmisión de la información de las protocélulas.
ORIGEN de los EUCARIOTAS
Las primeras células que aparecieron en la Tierra fueron las células procariotas hace unos 3500 millones de años. Procariota significa anterior al núcleo, es decir, no tienen el ADN encerrado en un compartimento membranoso. De hecho, estas células tienen una organización relativamente sencilla con una membrana que delimita un espacio interno donde se producen las reacciones químicas. Por fuera de la membrana tienen una cápsula y en ocasiones muestran prolongaciones como son los flagelos bacterianos que permiten la movilidad, y los pilis para el intercambio de material genético. Esta forma celular fue la única en los primeros años de la vida en la Tierra. Se conocen dos grandes grupos de procariotas: las bacterias y las arqueas.
1. ¿Cuándo?
La aparición de la célula eucariota fue un evento evolutivo que ocurrió hace unos 1500-2000 millones de años, es decir, unos 1500 millones de años después de que lo hicieran las primeras células procariotas. Su aparición supuso una transición evolutiva, es decir, fue algo nuevo y diferente a lo que había anteriormente y presentó suficientes novedades como para abrir nuevos caminos evolutivos hasta entonces inexplorados. Así, las células eucariotas llegaron a una complejidad morfológica y estructural no conocida hasta entonces, destacando un complejo sistema de compartimentos membranosos internos, incluidos el núcleo y el citoesqueleto. También fueron capaces de incorporar genomas completos (que dieron lugar a las mitocondrias y a los cloroplastos), descubrieron la reproducción sexual, y permitieron la aparición de algo desconocido hasta entonces: los organismos pluricelulares (cosa que ha ocurrido varias veces de forma independiente).
2. ¿De quién?
Esta cuestión no está resuelta todavía, pero se acepta que fue la consecuencia de la colaboración entre los dos tipos celulares que existían entonces: las arqueas y las bacterias. Se propone que los eucariotas son monofiléticos, es decir, todas las células eucariotas, incluyendo plantas, animales, hongos, algas y los eucariotas unicelulares, descienden de un único ancestro denominado LECA (last eukaryotic common ancestor). Por tanto, y mientras no se demuestre lo contrario, la célula eucariota sólo se inventó una vez por la evolución. Mediante el estudio comparado de genes se ha llegado a la conclusión de que LECA tenía un genoma tan complejo como los eucariotas actuales y probablemente era morfológica y estructuralmente parecida a los eucariotas actuales.
No cabe duda de que LECA se formó a partir de células procariotas, pero ¿a partir de cuál se formó, de las bacterias o de las arqueas? Hay un gran problema en esta elección. Las células eucariotas actuales parecen ser quimeras, en las que coexisten genes heredados de los dos tipos de procariotas. Unos genes trabajan en la traducción, transcripción y replicación de los genes (denominados genes informacionales) y están estrechamente relacionados con los de arqueas, mientras que los que están implicados en el metabolismo energético e intermediario, en la síntesis de componentes celulares como aminoácidos, lípidos y nucleótidos (denominados genes operacionales) son más parecidos a los genes bacterianos. Para complicar más la cosa, incluso aquellos genes de origen arqueano, no proceden de un solo grupo de arqueas, sino que son el legado de varios grupos. Sin embargo, numerosas evidencias tienden a colocar a los eucariotas en la rama de las arqueas (ver Figura 1).
Figura 1. En la imagen A se propone que los eucariotas se originaron a partir de un ancestro común compartido con las arquea. Este es el modelo 3D. En la imagen B se propone que LECA se formó directamente de una rama de los eucariotas. Éste es el modelo 2D. LECA (last eukaryotic common ancestor) es la célula de la que descienden todas las células eucariotas actuales.
Atendiendo a los estudios filogenéticos (comparación de secuencias de nucleótidos de algunos genes) se tiende a colocar a los eucariotas como descendientes de un grupo de arqueas. Actualmente se ha descubierto un grupo de arqueas denominado grupo Asgard, entre las que están las lokiarqueas, que son las procariotas más próximas evolutivamente a los eucariotas cuando se comparan secuencias de ciertos genes. Hay que tener cuidado con estas clasificaciones porque para ellas se utilizan los genes denominados informacionales (aquellos encargados de procesar el ADN y que se supone que han cambiado menos). Los que defienden este origen consideran que estos genes son los que se transmiten de “padres” a “hijos” directamente y no entre células no relacionadas, y por tanto los importantes a la hora de establecer relaciones evolutivas. Las lokiarqueas también tienen otros genes relacionados con el citoesqueleto y con la organización de las membranas internas homólogos a los de eucariotas. Curiosamente, todavía no se ha visto una lokiarquea, sino que su existencia se ha deducido por un estudio metagenómico, es decir, se cogió agua cerca de una fumarola y se aisló todo el ADN que contenía y posteriormente se estudió qué organismos había, analizando sólo el ADN. Se han encontrado en lugares anóxicos, lo que hace presuponer que las células vivían en ambientes sin oxígeno y que sólo gracias a la adquisición de las bacterias tolerantes al oxígeno (las futuras mitocondrias) pudieron colonizar ambientes más oxigenados. Al no saber cómo son morfológicamente no se pueden establecer similitudes con las células eucariotas en cuanto a tamaño o complejidad estructural.
Pero en realidad, en el núcleo de una eucariota hay 2 o 3 veces más genes de origen bacteriano que de arqueas. Hay otro problema adicional, las membranas de los eucariotas no tienen cadenas de isoprenoides en sus ácidos grasos ni enlaces tipo éter, ambos típicos de las membranas de las arqueas, y por tanto se puede decir que tienen una membrana más bacteriana. Por otra parte, hay quien aún sostiene que en realidad los eucariotas surgieron por una fusión directa entre una arquea y una bacteria y que los genes encargados de manipular el ADN fueron los de la arquea, mientras que los bacterianos se encargaron del metabolismo, incluida la síntesis de moléculas de membrana. Es decir, no hay una rama que parte de arqueas sino una rama completamente nueva creada a partir de dos ramas distantes.
3. ¿Cómo?
Hay un hecho clave en la aparición de LECA y es qué importancia tuvo la incorporación del antepasado de las mitocondrias. Hay autores que sugieren que esta incorporación fue la desencadenante y motor de la evolución hasta LECA, mientras que otros autores sugieren que la célula que engulló al antecesor bacteriano de las mitocondrias ya era muy compleja, tanto genómicamente como estructuralmente, y por tanto la endosimbiosis sólo fue un paso más en la evolución hasta LECA (ver Figura 2). Hay multitud de modelos que intentan explicar cómo ocurrió el proceso evolutivo que desembocó en LECA, pero hay dos líneas principales:
Figura 2. Modelos que explican la formación de la primera células euriota (LECA). Los colores de las membranas indican que tipo celular las controla (modificado de López-García y Moreira, 2015)
Modelo simbionte. Propone una fusión directa entre una arquea y una bacteria, y no existiría un protoeucariota como tal. Aquí habría sólo dos ramas principales de células iniciales, arqueas y bacterias. Los eucariotas serían una tercera rama surgida de la fusión de estas dos ramas. Hoy en día se han encontrado bacterias con endosimbiontes. Este evento de fusión dispararía el proceso de incremento de complejidad celular, y la bacteria terminaría siendo una mitocondria. En esta simbiosis las dos células se repartirían el funcionamiento celular: arquea el ADN y bacteria el metabolismo. Hay una variante de este modelo en la que la asociación entre bacterias y arqueas no tuvo por qué ser una incorporación de una célula dentro de otra en un momento determinado, sino que la asociación ocurrió a lo largo de mucho tiempo. Habría ocurrido transferencia lateral de genes de la bacteria a la arquea debido a que las condiciones ambientales favorecieron la proximidad física entre ambas. Se propone la teoría del hidrógeno en el que la bacteria produciría hidrógeno para el metabolismo de la arquea y la arquea produciría sustancias carbonadas que usaría la bacteria. Finalmente hubo una incorporación física de la bacteria dentro de la arquea, la cual ya tenía muchos genes bacterianos. Cómo se produjo esta inclusión no está claro. Casi se ha asumido que fue por fagocitosis, pero en realidad no hay ninguna evidencia experimental que apoye esta idea.
Modelo autógeno o endógeno. Existiría una célula protoeucariota que procedería de un ancestro común compartido con las arqueas. Esta célula habría evolucionado de manera independiente, adquiriendo la mayoría de las complejidades que aparecen en una célula eucariota actual, incluyendo endomembranas y citoesqueleto, pero aún no tendría a las mitocondrias. Tendría la capacidad fagocitar y en algún momento engulló a una alfaprotobacteria, que no fue digerida y pasó a vivir dentro de la protoeucariota. Con el tiempo los genes de la bacteria endosimbionte tomarían el control del metabolismo general, pero no de la manipulación del ADN. Sin embargo, no se han encontrado formas intermedias entre eucariotas y procariotas, y , sobre todo, no se han encontrado células eucariotas sin mitocondrias (aquellas células que no tienen mitocondrias tienen otros orgánulos derivados de éstas). Este modelo tiene además un gran hándicap, y es que para formar todo este sistema funcional de membranas se requieren muchas proteínas, y por tanto mucha energía, cosa que no parece que tal célula pudiera ser capaz de generar.
4. Endomembranas
Una de las características que distingue a las células eucariotas de las procariotas es su sistema de endomembranas, cuyo origen evolutivo todavía permanece sin aclarar. La idea tradicional es que tales compartimentos internos se produjeron por invaginación de la membrana plasmática de la procariota arquea, de modo que el interior del retículo o del aparato de Golgi es como si fuera homólogo al exterior celular. En relación con esto, y aceptando que la célula hospedadora fue un ancestro arqueano y la huésped fue un ancestro bacteriano, permanece la cuestión de cómo se cambiaron los lípidos de la membrana plasmática de origen arqueano (cadenas de isoprenoides y enlaces éter en los lípidos) por los de origen bacteriano actuales (lípidos sin isoprenos y sin enlaces tipo éster).
Los procariotas actuales no generan vesículas internas, es decir, no son capaces de hacer endocitosis, aunque sí pueden invaginar su membrana y crear cisternas membranosas internas que permanecen conectadas con la membrana plasmática. Sin embargo, tanto arqueas como bacterias pueden generar vesículas hacia el exterior, es decir, vesículas extracelulares. Es muy interesante que esta capacidad se mantiene en las mitocondrias actuales, las cuales son capaces de generar vesículas denominadas «vesículas derivadas de mitocondrias», las cuales quedan libre en el citosol.
Gould y colaboradores, (2016) proponen una aproximación nueva a cómo se pudo generar el sistema de endomembranas en las células eucariotas: sería el resultado de la fusión de vesículas liberadas por los ancestros de las mitocondrias que quedaron como endosimbiontes en el interior de una arquea. La generación y fusión de vesículas intracelulares explicaría el retículo endoplasmático/aparato de Golgi, y también el cambio de composición de la membrana por fusión de tales vesículas con la membrana arqueana. Todo este proceso no requeriría una gran innovación evolutiva. En este modelo, el interior del retículo endoplasmático no es homólogo al espacio extracelular sino a la región intermembranosa de la mitocondria.
Hay algunas evidencias que apoyan esta propuesta. Los contactos entre membranas del retículo endoplasmático y las mitocondriales son frecuentes, la N-glicosilación que acontece en el retículo endoplasmático también ocurre frecuentemente en bacterias, y en el espacio intermembranoso se almacena calcio, al igual que el interior del retículo. También hay indicios de que las mitocondrias pudieron ser las responsables de la formación de los peroxisomas por vesículas derivadas desde las membranas de las propias mitocondrias. Curiosamente, la envuelta nuclear, y por tanto el núcleo, sería más probable que se hubiese formado una vez que el retículo endoplasmático estuviese desarrollado, y no por fusión de las vesículas del endosimbionte.
La idea general es que la incorporación del ancestro bacteriano fue por fagocitosis, pero en este modelo el tráfico vesicular, y por tanto la fagocitosis, se desarrollaron después de la incorporación del ancestro de la mitocondria. La incorporación de un procariota dentro de otro procariota no es un gran problema porque hoy en día hay ejemplos de procariotas endosimbiontes de otras procariotas, que se han generado por mecanismos diferentes a la fagocitosis. Además, algunas proteínas necesarias para la formación de compartimentos de endocitosis o fagocitosis, como la dinamina, tienen homólogos en las bacterias, pero no en las arqueas.
Generar una célula con tal complejidad de membranas, con citoesqueleto, flagelos, etcétera, requeriría mucha energía. Por eso la teoría protoeucariota o arqueozoa de la generación autógena del sistema de endomembranas podría haber sido muy difícil sin una fuente de energía. Tal complejidad requiere una cantidad enorme de proteínas y aproximadamente el 75 % del gasto energético de una célula eucariota se consume en la síntesis de proteínas.
Un dato curioso es qué paso con el sistema de producción de ATP de la arquea. Las células procariotas son capaces de producir ATP en su membrana plasmática gracias a las proteínas ATPasa. Pero una vez incorporada una bacteria en su interior, con su propia ATPasa y con una capacidad de producción energética mucho mayor, la ATPasa arqueana pasó de la membrana plasmática al sistema de endomembranas y se estableció en el compartimento endosomal/lisosomal/vacuolar para funcionar de manera contraria: gastar ATP para transportar protones y acidificar tales compartimentos, permitiendo así la digestión intracelular.
ENDOSIMBIOSIS
Se cree que todos los organismos han evolucionado a partir de un tipo celular que apareció hace unos 3500 millones de años denominado LUCA (en inglés, Last Universal Common Ancestor). Esta célula debió ser sencilla, supuestamente semejante a los procariotas actuales. Sin embargo, la complejidad celular de algunas de estas primeras células aumentó, dando lugar a la aparición de las células eucariotas. Todas las células eucariotas actuales se cree que proceden de una de estas células iniciales, a la que se denomina LECA (en inglés, Last Eukaryotic Common Ancestor). Las células eucariotas actuales tienen compartimentos membranosos internos como el núcleo y diversos orgánulos como retículo endoplasmático, aparato de Golgi, endosomas, mitocondrias, cloroplastos, etcétera, además del citoesqueleto. Los primeros restos fósiles apuntan a que las células eucariotas estaban ya presentes hace unos 1500 millones de años, pero se cree que aparecieron mucho antes.
1. Definición
Mereschokovsky (1905, 1910) fue el primero en proponer que los cloroplastos son los descendientes de una célula procariota incorporada por una eucariota. A este proceso le llamó simbiogénesis, que derivó en el término endosimbiosis. Posteriormente se incluyeron a las mitocondrias también como resultado de un proceso de simbiogénesis. Así, tanto mitocondrias como cloroplastos constituyeron en el pasado formas libres de células primitivas procariotas. Estas células fueron incorporadas por otras células, evolucionaron y cambiaron con el tiempo, llegado hasta nuestros días transformadas en orgánulos celulares. Hoy en día se acepta que tanto mitocondrias como cloroplastos se originaron por endosimbiosis. Algunos autores han postulado que los peroxisomas, los cilios y los flagelos también se formaron por procesos de endosimbiosis, aunque hay poco soporte experimental.
2. Evidencias
La teoría de la endosimbiosis se basa en algunas semejanzas entre las bacterias actuales con las mitocondrias y los cloroplastos: ambos orgánulos tienen unas dimensiones parecidas a las bacterias, poseen hebras circulares de ADN en su interior y sus ribosomas son 70S, similares a los de las bacterias. Además, son capaces de replicarse de forma independiente en el interior celular. La doble membrana que presentan ya la tendrían cuando pasaron al interior de otra célula. En el caso de los cloroplastos se perdió la cubierta de peptidoglicano, pero las dos membranas también las poseía el ancestro. Mitocondrias y cloroplastos fueron inicialmente bacterias libres que se incorporaron o se internaron en otras células mayores (una arquea y una eucariota, respectivamente) y que llegaron a tal grado de dependencia que terminaron por perder su autonomía. Los antepasados de las mitocondrias podrían ser los antepasados de las alfa-proteobacterias actuales y los de los cloroplastos los antepasados de las cianobacterias actuales.
Los cloroplastos y las mitocondrias son muy diferentes a las cianobacterias y a las bacterias aerobias actuales. Por ejemplo, las cianobacterias actuales tienen unos 3000 genes, mientras que los cloroplastos actuales sólo poseen unos 100 o 200 genes. La pérdida de genes hace que los que quedan sólo codifican para un 10 % de sus proteínas. Esto es porque muchos de los genes cloroplastidiales han pasado al núcleo, el cuál se encarga de sintetizar muchos de los componentes que el cloroplasto necesita. Esto es un paso bastante complicado porque tales genes tienen que expresarse en un ambiente totalmente diferente y además, sus productos, las proteínas, deben viajar por el citosol, entrar en el cloroplasto y realizar su función en compartimentos concretos dentro del cloroplasto. La gran ventaja es que el núcleo celular coordina el funcionamiento y división de los cloroplastos. Un fenómeno similar ha ocurrido con las mitocondrias.
Se ha sugerido que la adquisición de la mitocondria como suministradora de mucha energía permitió aumentar la complejidad estructural de la célula eucariota. Algunos autores han propuesto que la principal propiedad que aportaron inicialmente los ancestros de las mitocondrias fue calor, lo que permitió a las células vivir en ambientes más fríos. La producción de ATP como principal elemento beneficioso para la célula llegó mucho más tarde durante la evolución. Las procariotas termófilas, que pueden vivir a temperaturas por encima de los 80ºC, son fundamentalmente arqueas. Se supone que las arqueas iniciales habrían vivido en esos ambientes que debieron ser muy frecuentes en aquellos tiempos en la superficie de la Tierra; luego podrían haber conquistado zonas más frías gracias a la capacidad de los ancestros de las mitocondrias de convertir el gradiente de protones generado en su membrana interna en calor, en vez de en ATP.
3. Proceso
La teoría de la endosimbiosis postula una primera fusión de procariotas (Figura 1). Hoy se favorece la idea de que fue entre una arquea y una bacteria. Esto se produjo probablemente tras un periodo de colaboración metabólica entre ambas células, es decir, hubo una simbiosis (no endosimbiosis todavía) previa a la incorporación de la bacteria en la arquea. Posteriormente a la incorporación, la célula desarrolló todo un sistema de orgánulos membranosos y un citoesqueleto, y la bacteria se convirtió en la mitocondria con el paso del tiempo. Así, tendríamos una célula similar a las eucariotas actuales. Posteriormente, hubo una segunda colonización de estas células eucariotas por parte de procariotas con clorofila, probablemente similares a las cianobacteras actuales, que con el tiempo se transformaron en los cloroplastos, resultando en las células fotosintéticas como las de los vegetales, que poseen tanto mitocondrias como cloroplastos. Es decir, se habrían producido dos endosimbiosis en serie, por lo que algunos autores hablan de la célula eucariota vegetal como una comunidad microbiana bien organizada.
Figura 1. Sucesos que supuestamente llevaron a la aparición de las mitocondrias y a los cloroplastos de las células eucariotas. Ocurrió mediante dos procesos independientes de endosimbiosis. Las células procariotas que se convirtieron en cloroplastos se cree que fueron similares a las cianobacterias actuales.
En la actualidad se conocen «comunidades celulares» todavía más complejas. Una endosimbiosis primaria (no confundir con la primera endosimbiosis) resulta de la asociación de una célula libre con otra célula, una de ellas termina dentro de la otra, lo que a la larga supone una gran alteración del ADN de la célula asimilada y del hospedador. Ambas células se han adaptado y evolucionado para mantener la endosimbiosis. Se conocen tres endosimbiosis primarias. Las más extendidas y que más impacto produjeron son la que dio lugar a las mitocondrias y la que dio lugar a los cloroplastos. Hay una tercera, que es una alfa cianobacteria qeu se incorporó en un eucariota unicelular denominado Paulinella chromatophora. Los descendientes de las tres endosimbiosis primarias tienen menos genes que una bacteria común y guardan los imprescindibles para su ciclo dentro del hospedador. Una endosimbiosis secundaria (no confundir con la segunda endosimbiosis) ocurrió cuando una célula eucariota con mitocondrias y cloroplastos se «zampó» a otra célula eucariota que ya contenía cloroplastos y mitocondrias (Figura 2). Con el tiempo la célula incorporada pasó a ser endosimbionte. La célula «ingerida» perdió el núcleo, o se atrofió, y sus cloroplastos pasaron a trabajar para la célula hospedadora y a depender de ella. Actualmente, se conocen tres sucesos independientes de endosimbiosis secundaria. La endosimbiosis terciaria ocurrió cuando una célula eucariota incorporó a otra eucariota que era resultado de una endosimbiosis secundaria. De todos estos casos hay ejemplos en la naturaleza.
Figura 2. Esquema del proceso de formación de una endosimbiosis primaria y otra secundaria.
Hoy en día se conocen muchos ejemplos de bacterias, pero ninguno de arqueas, que se localizan en células eucariotas a modo de simbiontes, incluso de bacterias dentro de arqueas, aunque no han llegado al grado de integración que observamos en mitocondrias y cloroplastos. Son diferentes caminos que se han explorado durante la evolución en la cooperación entre distintos tipos celulares. Cualquiera que sea el tipo, los simbiontes son capaces de proveer moléculas que el hospedador necesita. Por ejemplo, muchos invertebrados tienen bacterias que son intracelulares, las cuales llevan a cabo su ciclo de vida y pueden pasar a través de los gametos a su descendencia. Son simbiontes obligados que realizan su ciclo en el interior de las células del hospedador y se transmiten a la descendencia. Se han adaptado de tal manera que son inocuas para el hospedador, a veces son beneficiosas y otras necesarias. En realidad son infecciones que no producen daños importantes a los hospedadores, aunque usen la misma maquinaria que las bacterias patógenas para su reproducción. También hay endosimbiontes entre eucariotas. Por ejemplo, el paramecio Bursaria alberga en su interior una serie de algas del tipo Chlorella. Este protozoo busca siempre lugares bien iluminados gracias a su gran movilidad. El alga aprovecha esta alta intensidad de luz para realizar la fotosíntesis y de los productos resultantes se aprovecha el paramecio. Existen otros muchos ejemplos. Algunos simbiontes se denominan secundarios y no son permanentes, producen invasiones horizontales, es decir saltan entre individuos, su ADN no es tan grande como el de las bacterias libres, ni tan pequeño como el de otros simbiontes más integrados.
MATRIZ EXTRACELULAR
En el viaje por la célula que propuso C. de Duve (A guide tour of the living cell. Scientific American books, vol. 2, 1984) un citonauta de tamaño molecular, al dirigirse a una célula de un tejido animal, antes de toparse con la membrana plasmática, tendría la sensación de estar avanzando por una jungla de troncos, ramas y lianas. A esta maraña es a lo que denominamos matriz extracelular: un entramado de moléculas, sobre todo proteínas y carbohidratos, que se disponen en el espacio intercelular y que son sintetizadas y secretadas por las propias células.
La matriz extracelular es un invento de los organismos pluricelulares. Es esencial para estos organismos puesto que permite la adhesión de las células para formar tejidos. Pero con el tiempo ha adquirido muchas más funciones: mantiene la integridad y aporta las propiedades mecánicas a los tejidos (tanto en animales como en vegetales), mantiene e influye en la forma celular, permite la comunicación intercelular, forma sendas por las que se mueven las células, modula la diferenciación y la fisiología celular, secuestra factores de crecimiento, etcétera. Las propiedades que tienen algunos tejidos como resistencia, dureza, elasticidad, hidratación o propiedades ópticas, dependen de su matriz extracelular.
La importancia de la matriz celular para el correcto funcionamiento de los tejidos y órganos se pone de manifiesto en numerosas patologías donde las alteraciones de algún componente de la matriz extracelular llevan a severos desórdenes como las osteoartritis, fibrosis y cáncer.
La cantidad de matriz extracelular depende del tipo de tejido considerado. En los animales, hay algunos como el epitelial y el nervioso que tienen muy poca matriz extracelular, mientras que en otros, como el tejido conectivo propiamente dicho, el cartílago o el hueso, constituye la mayor parte del tejido (Figura 1). La composición molecular de la matriz extracelular es típica de cada tejido y sus componentes son renovados continuamente por las células que la producen. Esto supone que la matriz extracelular está en constante renovación.
Figura 1. En esta imagen se presentan ejemplos de distintos tipos de matrices extracelulares teñidas con diferentes colorantes. Los asteriscos señalan la matriz extracelular. A) Cartílago hialino. B) Matriz ósea compacta. C) Conectivo denso regular (tendón). D) Conectivo gelatinoso del cordón umbilical. E) Paredes celulares del sistema vascular de un tallo de una planta. F) Células epiteliales. Obsérvese que prácticamente no hay sustancia intercelular. G) Imagen de microscopía electrónica del tejido nervioso donde prácticamente no existe matriz extracelular.
Las principales macromoléculas que componen la matriz extracelular de los animales son: proteínas estructurales, fundamentalmente fibrosas, como el colágeno y la elastina, y componentes no fibrilares como los glicosaminoglicanos, proteoglicanos y glicoproteínas (Figura 2 y 3). Todos ellos interaccionan entre sí para formar el entramado funcional que es la matriz extracelular. En las plantas destacan la celulosa, hemicelulosa y lignina que forman la pared celular. Todas ellas se encuentran en un medio acuoso junto con otras moléculas de menor tamaño, además de iones. Es la cantidad, la proporción y el tipo de cada una de estas macromoléculas lo que distingue a unas matrices extracelulares de otras.
Figura 2. Esquema de las principales moléculas que aparecen en la matriz extracelular de un tejido conectivo de los animales. Figura 3. Imagen de microscopía electrónica de barrido de la matriz extracelular de la submucosa del digestivo de una rata. Las cintas largas son fibras de colágeno.
En los tejidos vegetales la pared celular se puede considerar, aunque no siempre hay acuerdo, como una matriz extracelular especializada con unas características muy diferentes a la de los tejidos animales. Su papel es crucial para dar rigidez a las células, y por extensión a la planta, es una barrera a la permeabilidad y protege frente a las agresiones mecánicas o frente a patógenos, entre otras funciones.
Las células interaccionan con la matriz celular mediante proteínas transmembrana, principalmente las integrinas, las cuales se adhieren o reconocen a moléculas de la matriz extracelular, y otros receptores de membrana, así como a través de moléculas liberadas desde la propia matriz extracelular. La matriz extracelular aporta a las células señales moleculares para la diferenciación, supervivencia, migración y proliferación, y para mantener la homeostasis del tejido donde se encuentran.
PROTEÍNAS ESTRUCTURALES
Índice de la página
1. Colágeno
– Organización
– Síntesis
2. Fibras elásticas
La matriz extracelular de los animales está formada principalmente por proteínas, glicosaminoglicanos, proteoglicanos y glicoproteínas, organizados en entramados diversos que constituyen las diferentes matrices extracelulares de los distintos tejidos. Las proteínas estructurales más abundantes son el colágeno y la elastina.
Colágeno
Se denomina colágeno a una familia de proteínas muy abundante en los animales, pudiendo representar del 25 al 30 % de todas las proteínas corporales. Tradicionalmente se ha usado el colágeno para fabricar pegamentos y colas, de ahí su nombre (del griego kola: pegamento). En los vertebrados hay unos 46 genes que sintetizan unas cadenas de aminoácidos denominadas cadenas alfa, las cuales se asocian de tres en tres para formar hasta 28 tipos de moléculas de colágeno diferentes (ver tabla). La combinación de cadenas alfa puede ser homotípica (todas las cadenas iguales) o heterotípica (cadenas diferentes). La principal misión del colágeno es crear un armazón que hace de sostén a los tejidos y que resiste las fuerzas de tensión mecánica. Actúa como las barras de acero que refuerzan el hormigón en los edificios. La organización de las moléculas de colágeno en estructuras macromoleculares tridimensionales es variada, pudiendo formar haces, matrices, etcétera. Las células se «agarran» a las moléculas de colágeno mediante diversas proteínas de adhesión como las integrinas, inmunoglobulinas, anexinas, etcétera. Otras moléculas de la matriz extracelular interaccionan también con las moléculas de colágeno como proteoglicanos y glicoproteínas, trabando más todo la estructura.
Organización
Las moléculas de colágeno se caracterizan por:
a) Una composición poco frecuente de aminoácidos. En las moléculas de colágeno abunda el aminoácido glicina, y hay otros menos comunes como la hidroxilisina y la hidroxiprolina. La glicina se repite cada 3 aminoácidos (…-Gly – x – y – Gly – x – y -…), donde x e y suelen ser hidroxilisina e hidroxiprolina, respectivamente. Esta secuencia repetida con la glicina, y debido al pequeño tamaño de este aminoácido, es la que permite la disposición en hélice levógira de las cadenas polipetídicas formando así las cadenas alfa.
Figura 1. Fibras de colágeno en la matriz extracelular del tubo digestivo. Microscopía electrónica de barrido.
Figura 2. Fibras de colágeno de la dermis. Tinción: tricrómico de Masson.
Figura 3. Fibras de colágeno en la matriz extracelular del tubo digestivo. Microscopía electrónica de transmisión.
b) Pueden organizarse formando fibras, mallas o especializarse en formar uniones entre moléculas. Todo ello depende de la composición química de sus subunidades alfa y de los tipos de subunidades que lo formen (ver tabla).
Forman fibras. Son las más abundantes de todas las formas de colágeno y están formadas por repeticiones alineadas de moléculas de colágeno repetidas (Figuras 1, 2 y 3). De los colágenos que forman fibras el más frecuente es el tipo I, que abunda en huesos, cartílago y piel, y que representa el 90 % de todo el colágeno del organismo. Otros tipos abundantes son el II, presente en el cartílago hialino, y el III, que abunda en la piel y en los vasos sanguíneos. Igual que hay moléculas heterotípicas (cadenas alfa diferentes), también hay fibras heterotípicas formadas por más de un tipo de colágeno. Por ejemplo, hay fibras de colágeno I que contiene también colágeno tipo III y V.
Forman mallas. Estos tipos de colágeno suelen organizarse en entramados moleculares que forman láminas. Se encuentran rodeando los órganos, rodeando células como las musculares, o formando la base de los epitelios. Entre éstos se encuentra el colágeno tipo IV que abunda en la lámina basal, localizada entre el epitelio y el tejido conectivo.
Se organizan en forma de pequeños filamentos arrosariados que se distribuyen en un entramado que ayuda a la cohesión de la matriz extracelular. Destaca el colágeno tipo VI.
Están asociados a otras fibras de colágeno y modulan la disposición, organización e interacción de dichas fibras con otros elementos del citoesqueleto. Por ejemplo, el colágeno tipo IX forma uniones entre los glicosaminoglicanos y las fibras de colágeno tipo II.
También existen moléculas de colágeno que poseen secuencias de aminoácidos hidrofóbicos y que se encuentran como moléculas transmembrana. Es el caso del colágeno tipo XIII y el tipo XVII. Estos colágenos operan como receptores y están relacionados con la adhesión y movilidad. El colágeno tipo XVII forma parte de la estructura de los hemidesmosomas.
Síntesis
Independientemente del tipo, la síntesis de las moléculas de colágeno ocurre en forma de precursor (Figura 4). El colágeno se sintetiza en el interior celular en forma de procolágeno. En primer lugar se sintetizan las cadenas alfa inmaduras en el retículo endoplasmático, donde son modificadas. Las prolinas y lisinas son hidroxiladas para dar hidroxiprolinas e hidroxilisinas, pudiendo representar ambas hasta el 20 % de la molécula de colágeno. También se lleva a cabo glicosidación (del tipo O-glicosidación). En este momento se asocian las cadenas alfa de 3 en 3 gracias a puentes de hidrógeno y a puentes disulfuro, para formar las moléculas de procolágeno. Éstas son reconocidas por receptores transmembrana y empaquetado en vesículas recubiertas por COPII. Estas vesículas, de unos 500 nm de diámetro, han de ser diferentes puesto que las moléculas de procolágeno son como varillas rígidas de unos 300 nm (las vesículas típicas COPII miden entre 60 y 90 nm). El procolágeno pasa por el aparato de Golgi, no se sabe muy bien cómo, desde donde es exocitado al exterior celular. Es destacable que algunas células pueden seleccionar el dominio celular donde se liberará un determinado tipo de colágeno. Independientemente de esto, durante, o tras la liberación, sufre una acción enzimática que elimina una secuencias terminales de cada cadena alfa por metaloproteinasas específicas, transformando el procolágeno en colágeno. Estas secuencias terminales impedían que el procolágeno se ensamble espontáneamente en el interior celular. Antes se pensaba que todo el procesamiento del procolágeno y el ensamblado de las fibrillas era extracelular, pero parece que empieza antes de su completa exocitosis en compartimentos entre el aparato de Golgi y la membrana plasmática.
Figura 4. Esquema de la síntesis de las fibras de colágeno.
Las moléculas de colágeno, sin cadenas terminales, se ensamblan automáticamente para formar las fibrillas de colágeno, que a su vez se unen para formar las fibras de colágeno (Figuras 4 y 5). La formación de las fibras de colágeno, sin embargo, parece estar controlada por la participación de los colágenos tipo V y XI. En concreto el colágeno tipo V parece imprescindible para la formación final de las fibras de colágeno ya que si se elimina no se observan tales fibras. La forma y el crecimiento de las fibras de colágeno se ven afectados por otras moléculas como algunos proteoglicanos como la decorina, fibromodulina y limicano. En la fase final de ensamblaje, y para dar estabilidad a la fibra, se forman enlaces covalentes por enzimas como la lisil oxidasa.
Figura 5. Imagen obtenida con un microscopio electrónico de transmisión a partir de tejido conectivo de un invertebrado marino, la oreja de mar. Con los asteriscos negros se indica el colágeno ya ensamblado en el exterior celular, mientras que con los asteriscos blancos las grandes vesículas intracelulares llenas de moléculas de procolágeno. La flecha blanca indica un posible punto de liberación de las moléculas de procolágeno al espacio extracelular.
El colágeno se sintetiza principalmente por fibroblastos, miofibroblastos, osteoblastos y condrocitos. Aunque, en general, todas las células de un tejido pueden contribuir a la síntesis de matriz extracelular. Por ejemplo, algunas moléculas de colágeno son también sintetizadas por otros tipos celulares tales como las células epiteliales.
Fibras elásticas
Figura 6. Fibras elásticas del tejido conectivo.
Figura 7. Fibras elásticas de la pared de una arteria elástica.
La elastina es un polímero proteico insoluble abundante en muchas matrices extracelulares y aparece como un componente de las denominadas fibras elásticas (Figuras 6 y 7). El principal componente de las fibras eláticas es la tropoelastina. Esta proteína se libera a la matriz extracelular y forma agregados amorfos entre los que se establecen enlaces covalentes. La tropoelastina posee una larga cadena de aminoácidos en la que hay numerosas secuencias con aminoácidos hidrófobos, separadas por otras secuencias que contienen parejas de glicinas y otros aminoácidos pequeños como la lisina. Esta composición de aminoácidos es la que confiere las propiedades elásticas, puesto que los aminoácidos hidrófobos permiten la disposición en estructuras arrolladas y la lisina la formación de α-hélices. De manera que en ambientes acuosos la tropoelastina adopta un disposición globular, pero que se puede estirar cuando se somete a fuerzas mecánicas. Cuando estas fuerzas desaparecen la proteína se vuelve de nuevo globular. Los aminoácidos no hidrófobos son los puntos donde se enlazan dos moléculas de tropelastina próximas para formar el polímero de elastina. El alto contenido en aminoácidos hidrófobos hace a la tropolastina una de las proteínas más resistentes y durables de la matriz extracelular. La tropoelastina parece ser una invención de los vertebrados, puesto que no se ha encontrado en invertebrados.
Al contrario que las fibras de colágeno, las fibras elásticas tienen la capacidad de estirarse en respuesta a las tensiones mecánicas y de contraerse para recuperar su longitud inicial en reposo gracias a las propiedades de la tropoelastina. Una fibra elástica está formada por el material amorfo de tropoelastina, que representa el 90 %, y por una cubierta formada por las denominadas microfibrillas de fibrilina y por otras glicoproteínas y proteoglicanos en menor proporción (Figura 8). La elasticidad de nuestros tejidos depende de las fibras elásticas. Estas fibras se encuentran sobre todo en la dermis, en las paredes de las arterias, en el cartílago elástico y en el tejido conectivo de los pulmones. La concentración de fibras elásticas varía entre tejidos. Por ejemplo, llega hasta el 70 % de la matriz extracelular en los ligamentos. Con el tiempo, la cantidad de fibras elásticas disminuye en los tejidos y los órganos y estructuras se vuelven más rígidas, como por ejemplo los vasos sanguíneos. Además de la elasticidad, otras funciones de las fibras elásticas son aportar sostén a los tejidos o regular la actividad de los factores de crecimiento TGF-β mediado por la fibrilina.
Figura 8. Esquema de una porción de una fibra de elastina. Las moléculas de elastina están unidas entre sí mediante enlaces entre las regiones ricas en el aminoácido lisina (Modificado de Kielty 2007).
GLÚCIDOS. PROTEOGLICANOS.
La matriz extracelular está formada por proteínas y glúcidos en un medio acuoso. En los tejidos animales las proteínas colágeno, las fibras elásticas y otras moléculas presentes en la matriz extracelular se encuentran embebidas en un medio compuesto fundamentalmente por polímeros no ramificados de glúcidos y agua. Los glúcidos corresponden sobre todo al tipo de los denominados glicosaminoglicanos o mucopolisacáridos. En las paredes celulares de las células vegetales no existen glicosaminoglicanos sino sobre todo celulosa, que es un polímero de glucosa.
1. Glicosaminoglicanos
Figura 1. Composición de los principales grupos de glicosaminoglicanos (Modificado de Lamoureux, 2007).
Son polímeros no ramificados de azúcares que pueden formar cadenas muy largas. Están formados por repeticiones de parejas de monosacáridos donde uno de los azúcares tiene un grupo amino (N-acetilgalactosamina o N-acetilglucosamina) y el otro es normalmente la galactosa o el ácido glucurónico. Estos azúcares poseen grupos carboxilo (COO–) y pueden tener grupos sulfatos (SO3–), cuyas cargas negativas permiten una fuerte y abundante asociación con moléculas de agua, aportando una gran hidratación a la matriz extracelular (Figura 1). Los glicosaminoclicanos son moléculas poco flexibles por lo que ocupan un gran volumen y gracias a su fuerte hidratación hacen que la matriz extracelular se comporte como un gel. Esto permite que los tejidos que poseen una alta proporción de glicosaminoglicanos puedan resistir fuertes presiones mecánicas y además favorece una alta tasa de difusión de sustancias entre las célula. Los tipos más comunes de glicosaminoglicanos son el ácido hialurónico y los glicosaminoglicanos que forman proteoglicanos: condroitín sulfato, dermatán sulfato, queratán sulfato y heparán sulfato.
Ácido hialurónico
ácido hialurónico
El ácido hialurónico o hialuronato es un caso especial de glicosaminoglicano puesto que no forma enlaces covalentes con otras moléculas de la matriz extracelular, se sintetiza por enzimas localizados en la membrana plasmática y no posee grupos sulfatos. Está compuesto por parejas de azúcares formadas por la N-acetil-D-glucosamina y el ácido D-glucurónico, que pueden llegar hasta las 20.000 repeticiones. Se suele asociar con las moléculas de colágeno o a proteoglicanos, confiriendo a la matriz extracelular elasticidad, resistencia y lubricación. Su función es muy importante durante el desarrollo o en lugares del organismo donde se produce una fuerte proliferación celular puesto que facilita el desplazamiento celular y difusión molecular. Al ser una molécula grande y poco flexible ocupa un volumen grande dejando muchos espacios. También aparece en aquellos lugares donde se produce una fuerte fricción como en el cartílago de las articulaciones.
Otros
Los demás tipos de glicosaminoglicanos están sulfatados y unidos covalentemente a cadenas de aminoácidos formando los denominados proteoglicanos (ver más abajo). El condroitín sulfato es un glicosaminoglicano sulfatado compuesto por repeticiones de parejas de N-acetil-galactosamina y ácido glucurónico. Es abundante en el cartílago y tejido nervioso. El dermatán sulfato está formado por ácido glucurónico o ácido idurónico más N-acetil-galactosamina. El queratán sulfato es un polímero de parejas de N-acetil-glucosamina más galactosa, con los azúcares mostrando distinto tipo de sulfatación. El heparán sulfato lo producen la mayoría de las células. Es un componente esencial de la lámina basal, donde constituye la mayor parte de ésta. Todos estos glicosaminoglicanos tienen muchas cargas negativas, lo que hace que se asocien fácilmente con otras moléculas como factores de crecimiento o quimiocinas, lamininas y fibronectinas. El derivado del heparán sulfato, la heparina, sólo la producen los mastocitos. La heparina es un antitrombótico muy usado en medicina como anticoagulante. Está formado por dímeros de N-acetil-glucosamina más ácido glucurónico o ácido idurónico, igual que el hialuronato, pero con distintos tipos de enlaces entre azúcares, los cuales son sulfatados. Todos estos glicosaminoglicanos están unidos covalentemente a cadenas de aminoácidos formando los denominados proteoglicanos.
2. Proteoglicanos
Un proteoglicano es una molécula compuesta por la unión covalente entre una cadena de aminoácidos y uno o varios glicosaminoglicanos sulfatados (Figura 2). Los glicosaminoglicanos se unen a los péptidos a través de residuos del aminoácido serina. Es una familia de moléculas ubicua en los tejidos animales y son elementos esenciales del espacio pericelular. Virtualmente todas las células son capaces de sintetizar proteoglicanos y secretarlos, dejarlos en sus membranas o almacenarlos en gránulos. Una célula es capaz de sintetizar varios tipos de proteoglicanos. Los proteoglicanos son sintetizados en el interior celular. La parte proteica se sintetiza en el retículo endoplasmático, donde también se inicia la adición de glúcidos. Sin embargo, la elongación de las cadenas de repeticiones de glicosaminoglicanos y la sulfatación se produce en el lado trans del aparato de Golgi. La mayoría de los proteoglicanos son exocitados al espacio intercelular, pero algunos formarán parte de la membrana plasmática gracias a que su parte proteica contiene secuencias de aminoácidos hidrófobos que se insertan entre las cadenas de ácidos grasos de la membrana.
Figura 2. Esquema de un proteoglicano, el agrecano (Modificado de Lamoureux, 2007).
Los proteoglicanos se diferencian sobre todo en la secuencia y en la longitud de la cadena de aminoácidos (desde 100 a 4000 aminoácidos). Relativamente pocas proteínas (o cadenas de polipéptidos) pueden tener unidos glicosaminoglicanos covalentemente (unas 17 para el heparán sulfato, 20 para condroitín/dermatán y 8 para el queratán). Ello no impide que se puedan producir una gran diversidad de proteoglicanos. Por ejemplo, se diferencian en el número y en el tipo de moléculas de glicosaminoglicanos que tiene unidos. Así, la decorina tiene una sola molécula mientras que el agrecano contiene más de 200. También el grado de sulfatación y en la distribución de los grupos sulfatos en la moléculas de glicosoaminoglicano. Por ejemplo, el grado de sulfatación varía con la edad, tejido y patología.
Los proteoglicanos se pueden agrupar en varias familias. Los lecticanos poseen una parte proteica con un extremo amino globular que interactúa con el hialuronato, un dominio central de coindritín sulfato y otro dominio terminal que actúa como una lectina tipo C. Posee sobre todo condroitín sulfato y, a veces, queratán sulfato. Son miembros de este grupo el agrecano, versicano, neurocano y brevicano. El agrecano abunda en el cartílago, el brevicano y neurocano se expresan en tejido nervioso, y el versicano en tejido conjuntivo. El grupo denominado SLRP se caracteriza por tener muchas repeticiones del aminoácido leucina y está unido a condroitín o dermatán sulfato y a queratán sulfato. Miembros de este grupo son la decorina, biblicano, fibromodulina o el queratocano. Una función de los proteoglicanos SLRP es estabilizar la fibras de colágeno. Un grupo importante es el formado por los proteglicanos que contienen heparán sulfato. Pueden encontrarse de tres maneras: unido a la superficie celular a través del receptor CD44v3 (como los sindecanos), unido covalentemente a los glucosil-fosfatidilinositoles de la membrana (como los glipicanos) o libres en la matriz (tales como la agrina, el colágeno tipo XVIII y el perlecán). Hay un grupo heterogéneo de proteínas entre las que se encuentran el receptor CD44, proteína precursora amiloide y algunos colágenos (IX, XII,XIV y XVIII) que pueden tener unidas o no moléculas de glicosaminoglicanos sulfatados. Es decir, serían proteoglicanos a «tiempo parcial».
La actividad biológica de los proteoglicanos depende en gran medida del peso molecular, monosacáridos que poseen y las uniones entre los disacáridos de los glicosaminoglicanos. Todos los glicosaminoglicanos tienen cargas negativas, pero la cantidad de cargas es una característica fundamental para su función. Esta carga negativa se asocia a la cantidad de grupos sulfato que posean, excepto en el ácido hialurónico, donde las cargas se deben al ácido glucorónico. La variacion en la posición de los grupos sulfatos hace que una misma cadena de azúcares pueda convertirse en cientos de moléculas diferentes. Estas funciones son: hidratación, resistencia a presiones mecánicas, lubricantes, afectan a la diferenciación, la movilidad y la fisiología celular, etcétera. Su acción mecánica es esencial en los cartílagos y en las articulaciones. Pero además son puntos de anclaje de las células a la matriz extracelular que les rodea, bien por su acción directa al ser moléculas integrales de la membrana plasmática, porque forman uniones con fosfolípidos de la membrana o porque son reconocidos por proteínas de adhesión como las integrinas, presentes en las membranas plasmáticas. Es interesante resaltar que los proteoglicanos sulfatados son escasos en los nichos de las células madres.
Los proteoglicanos pueden crear una barrera pericelular física y química. Las redes perineuronales son porciones de matriz extracelular que rodean el soma y dendritas proximales de neuronas del sistema nervioso central, donde podrían participar en la plasticidad, formación de nuevas sinapsis, etcétera. Su déficit está relacionado con ciertas patologías. Estas redes son inhibidores físicos que limitan la plasticidad neuronal.
Los glicosaminoglicanos sulfatados son también lugares de secuestro de proteínas cargadas, para su posterior uso. En algunas ocasiones la interacción de los glicosaminoglicanos con las proteínas provocan un cambio de conformación de éstas últimas, llevando a su activación o inactivación. Un ejemplo, es la heparina, la cual activa a la molécula antitrombina para que una a la trombina y factor X, con lo que se evita la coagulación sanguínea. Otro ejemplo es la fibronectina, la cual tras interactuar con glicosaminoglicanos cambia su conformación y expone más dominios con los que interactuar con otras moléculas de la matriz extracelular. Otra función de los glicosaminoglicanos es actuar como correceptores, de manera que la unión al receptor aumenta la afinidad por el ligando.
3. Celulosa
La celulosa es el principal componente de las paredes vegetales, la matriz extracelular de las plantas. Aunque algunos autores consideran que la pared celular de las plantas no se puede incluir en la categoría de matriz extracelular por sus características propias, nosotros la consideraremos como un tipo muy especializado de matriz extracelular. La celulosa es una estructura para cristalina compuesta de un polisacárido formado por monómeros de glucosa (más de 500 por molécula) unidos mediante enlaces tipo β(1-4) (Figura 3, y figura ). Las moléculas de glucosa se asocian entre sí mediante enlaces de hidrógeno y fuerzas de Van der Waals para formar estructuras denominadas microfibrillas de celulosa, formadas por unas 50 moléculas de celulosa orientadas con la misma polaridad. Las microfibrillas se asocian entre sí mediante enlaces formados entre ellas y por otros glúcidos, principalmente la hemicelulosa y las pectinas, que resultan en las fibrillas y fibras de celulosa, visibles al microscopio óptico.
Figura 3. Organización de las moléculas de celulosa y la interacción entre ellas mediante puentes de hidrógeno (líneas en rojo). Pared celular
Al igual que ocurre con el hialuronato, la celulosa se sintetiza en la membrana celular gracias a la acción de la celulosa sintasa, una proteína transmembrana con una secuencia de aminoácidos que cruzan 8 veces la membrana celular (Figura 4). Esta enzima recoge las unidades de glucosa desde el citosol, les hace cruzar la membrana y las enlaza en el exterior celular. Hasta 36 enzimas celulosa sintasa se unen en un punto de la membrana para formar el denominado complejo de celulosa sintasa, el cual puede sintetizar hasta 36 moléculas de glucosa simultáneamente, las cuales se van asociando en microfibrillas de celulosa conforme se van sintetizando.
Figura 4. Síntesis y orientación de las fibrillas de celulosa guiada por los microtúbulos. Los complejos sintetizadores de celulosa se desplazan a medida que van sintetizando la celulosa siguiendo los trayectos marcados por los microtúbulos (Modificado de McFarland et al., 2014). Un aspecto interesante de la síntesis de celulosa es la orientación de las moléculas que se van sintetizando, ya que determina la orientación de las microfibrillas y posteriormente de las fibras de celulosa. El crecimiento de las plantas es sobre todo por crecimiento del tamaño celular, el cual se produce por presiones hidrostáticas. Una célula crece hacia donde menos resistencia encuentre, lo cual depende de la resistencia que oponga la pared celular. La orientación de la deposición de las moléculas de celulosa condiciona esa resistencia, de manera que hay menor resistencia en la dirección perpendicular a las fibras de celulosa. La orientación de las fibras de celulosa está condicionada por los espacios por los que se puede mover la celulosa sintasa, que a su vez depende de la orientación de los microtúbulos corticales que se localizan justo debajo de la membrana plasmática, en el citosol (Figura 4). Estos microtúbulos son barreras que no pueden ser cruzadas por las enzimas sintasas de la celulosa. Estas enzimas se desplazan por las membrana a medida que van sintetizando fibrillas de celulosa pero sólo hacia donde les permiten los microtúbulos. Otros factores extracelulares e intracelulares pueden condicionar también la dirección del movimiento de estos complejos enzimáticos. De esta manera la célula puede controlar la orientación de las fibras de celulosa y por tanto su dirección de crecimiento, por ejemplo para que una estructura vegetal, tallos u hojas, crezcan hacia una fuente de luz. Además, de la orientación de las fibras de celulosa, la pared celular se hace más blanda en determinados lugares mediante la modificación química de la pectina (otro componente de la matriz extracelular) y por acidificación, y es en estos lugares donde también se encuentra menos resistencia y por tanto por donde crece la célula.
GLICOPROTEÍNAS
Las células están adheridas a la matriz extracelular, la cual, a su vez, forma un entramado cohesionado gracias a la interacción entre las moléculas que la forman. La mayoría de estas uniones en la matriz extracelular son entre proteínas, pero también entre proteínas y azúcares. Hay tres tipos de uniones que dan cohesión a los tejidos: uniones entre moléculas de la matriz extracelular, uniones entre las células y la matriz extracelular y uniones entre células contiguas. Los dos últimos tipos se verán en el apartado de membranas puesto que participan moléculas transmembrana.
Las moléculas que favorecen que la matriz extracelular sea un entramado cohesionado son principlamente glicoproteínas, aunque no es esa sola su función. Las glicoproteínas establecen puentes entre las moléculas estructurales de la matriz extracelular, y entre ellas y las células. Tienen múltiplesdominios de unión que reconocen y unen una gran variedad de moléculas, lo que les permite formar dichos entramados. Entre estas glicoproteínas destacan las fibronectinas, las lamininas y las tenascinas.
1. Fibronectina
Figura 1. Esquema de una molécula de fibronectina. Está formada por dos cadenas de aminoácidos unidas por la zona próxima al extrremo carboxilo por puentes disulfuro. Se indican los dominios de la proteína que interaccionan con otras moléculas produciendo adhesión. (Modificado de Pankov 2002)
Las fibronectinas son glicoproteínas formadas por dos cadenas de polipéptidos unidos por uniones disulfuro (Figura 1). Poseen dominios en su estructura que permiten unirse al colágeno, a ciertos proteoglicanos, a glicosaminoglicanos, a la fibrina, a la heparina y a proteínas de la membrana plasmática celular como las integrinas. Por tanto establecen uniones entre moléculas de la matriz extracelular y entre moléculas de las células con la matriz extracelular. Es una de las principales responsables de trabar la matriz extracelular. Una vez liberadas a la matriz extracelular pueden interactuar entre sí para formar largas fibras en torno a la célula. Las moléculas de fibronectina se encuentran en prácticamente todos los tejidos y pueden aparecer formando fibras insolubles en los tejidos conectivos o solubles en el plasma de los fluidos corporales, como la sangre. Tienen un papel muy importante durante el desarrollo embrionario creando sendas por las que pueden migrar las células de un lugar a otro del embrión, y también es abundante en tejidos en procesos de remodelación.
2. Laminina
Figura 2. Esquema de una molécula de laminina. Está formada por tres cadenas de aminoácidos unidas. Se indican los dominios de la proteína que interaccionan con otras moléculas produciendo adhesión. (Modificado de Mouw et al., 2014)
Las lamininas son uno de los principales componentes de la lámina basal. Su estructura proteica está formada por tres cadenas de aminoácidos altamente glicosidadas unidas por puentes disulfuro (Figura 2). Estas cadenas son alfa, beta y gamma. Existen 5 tipos de cadenas alfa, 3 betas y 3 gamma, las cuales se combinan entre sí para formar diferentes tipos de lamininas, aunque no todas las combinaciones son posibles ya que se han aislado sólo 16 formas de laminina en humanos. La laminina es sintetizada por células epiteliales, musculares, neuronas y células de la médula ósea. Epitelio y células musculares depositan la laminina principalmente en las láminas basales que las separan del tejido conectivo. Aparte de su función estructural las lamininas afectan a la diferenciación y comportamiento celular gracias a que son reconocidas por las integrinas. Por ello, defectos en las lamininas suelen conllevar procesos patológicos. Son de las primeras glicoproteínas de la matriz extracelular que aparecen en el embrión.
3. Glicoproteínas matricelulares
En torno a los años 90 del siglo pasado se comprobó que algunas proteínas de la matriz extracelular producían pérdida de adhesión de las células más que aumentarla y además cuando se bloqueaba el gen para estas proteínas los animales no parecían notarlo. Son proteínas que, aunque tienen lugares de reconocimiento para el colágeno, fibronectina y receptores celulares de superficie, su función no es eminentemente estructural. Estas glicoproteínas se caracterizan por ser secretadas de forma específica temporal y espacialmente por una gran diversidad de células, se asocian pero no forman parte necesariamente de la parte estructural de la matriz extracelular, pueden disminuir la adhesión de las células en ciertas circunstancias, están siempre presentes cuando los tejidos se están remodelando, y son abundantes durante el periodo embrionario. Es decir, son proteínas temporales más que constitutivas en la matriz extracelular.
La función principal de las proteínas matricelulares es modificar la actividad celular y favorecer la remodelación de la matrix extracelular. Esto ocurre durante el periodo embrionario y en la remodelación de los tejidos por causas naturales o patológicas. Son capaces de unirse a factores de crecimiento y modifican los efectos de estos factores en las células. Algunas de estas moléculas incluso tienen efectos en las células durante su fase intracelular, antes de ser liberadas a la matrix extracelular. Proteínas matricelulares son, por ejemplo, la tenascina, fibulina, tromboespondinam, osteopontina y la proteína SPARC.
Tenascinas
Figura 3. Esquema de la molécula de tenascina. Es una estructura modular donde se repite un polipéptido 6 veces, todos conectados entre sí. Cada color representa un dominio molecular.
Las tenascinas forman una familia de glicoproteínas de gran tamaño presentes en la matrix extracelular con una estructura molecular haxamérica modular (Figura 3). Se producen diferentes tipos de tenascina por maduración alternativa de su ARN mensajero. Hay varios tipos. La tenascina-C fue el primer tipo descubierto y se libera a la matrix extracelular de tendones, huesos y cartílago durante el desarrollo embrionario, pero también en otros tejidos del embrión. La tenascina-C no se expresa mucho en tejidos adultos pero sí cuando ocurren daños tisulares como en los infartos cardiaco. La tenascina-R es abundante en el sistema nervioso, tanto durante el desarrollo como en el adulto. La tenascina-X es abundante en el tejido conectivo y se expresa mucho en músculos que tienen que hacer esfuerzos, como en los deportistas. También se han descrito las tenascinas Y y W. La tenascina-Y es el equivalente aviar de la tenascina-X. Como otras glicoproteínas, la tenascina cambia la cohesión de la matriz extracelular puesto que puede establecer enlaces con integrinas, fibronectinas, colágeno y proteoglicanos. En condiciones normales, cada tipo de tenascina se expresa en lugares concretos del organismos, que pueden cambiar según el estado de desarrollo, pero también pueden inducirse cuando hay reparación de tejidos, durante procesos tumorales o patológicos.
Osteopontina
La osteopontina está presente en el hueso, relacionada con la mineralización y remodelación ósea, y en el riñón. En menos concentración aparece en el cartílago donde reconoce a proteoglicanos.
Fibulina
Las fibulina son un grupo de 7 glicoproteínas secretadas en muchos tejidos asociadas a la lámina basal, fibras elásticas y otros componentes de la matriz extracelular. Cada una de ellas se expresa de manera diferencial en diferentes del tejido y en diferentes etapas durante el desarrollo. La fibulina 5 es importante para la formación de las fibras elástica gracias a que puede unirse a la tropoelastina. Las fibulinas, además de su función en el entramado de la matriz extracelular afectan a la fisiología de las células. Así, estas proteínas se comportan como matricelulares y estructurales.
4. Metaloproteinasas. Remodelación de la matriz extracelular.
La matriz extracelular de los animales está en constante remodelación mediante la degradación de componentes y la producción de otros nuevos por parte de las células. La degradación de la matriz extracelular la llevan a cabo enzimas como las metaloproteinasas. Son enzimas que se asocian a la cara externa de la membrana plasmática (son secretadas) o forman parte integral de ella, siempre con su centro activo localizado extracelularmente. Inicialmente se producen en forma inactiva o prometaloproteinasas y para su activación es necesaria una proteolisis en su estructura, llevada a cabo por otras enzimas asociadas a la membrana plasmática. Existen unas 29 metaloproteinasas diferentes en mamíferos, cada una de las cuales tiene apetencia por distintos componentes de la matriz extracelular. Así, aunque no son totalmente específicas, se clasifican en colagenasas, gelatinasas, estromalisinas, matrilisinas, metaloproteinasas transmembrana y unidas a proteínas de membrana (Fgiura 4), y un grupo adicional heterogéneo. Cada grupo tiene un grupo de sustratos sobre el que tienen más apetencia. Las metaloproteinasas son capaces de degradar también receptores de membrana, proteínas de adhesión, citocinas, etcétera. Curiosamente se han encontrado metaloproteinasas en el núcleo.
Figura 4. Principales grupos de metaloproteinasas, sus miembros (tipos), y los sustratos sobre los que actúan. El grupo heterogéneo no se ha incluido. (Adaptado de Berg et al., 2019)
Las metaloproteinasas no sólo intervienen en el mantenimiento de la matriz extracelular sino que son cruciales en procesos como el desarrollo, remodelación de la matriz extracelular inducida por estímulos, por ejemplo, hormonas, o en procesos patológicos tales como la inflamación, reparación de tejidos o metástasis tumoral. Otro papel importante de la metaloproteasas es liberar moléculas que se encuentran ancladas en la matriz extracelular, las cuales actúan como señal sobre las celulas, y que se vuelven solubles tras la degradación de la matriz extracelular. Estas enzimas no sólo son producidas por los fibroblastos sino que también las sintetizan las células epiteliales, condrocitos, osteoclastos y leucocitos, además de células malignas como las tumorales.
ciclos gaseosos sedimentarios e hidrológicos son procesos fundamentales en la Hidrología, ciencia dedicada al estudio del agua…
La fotosíntesis es el proceso mediante el cual las plantas transforman materia inorgánica en materia orgánica.
La biosfera es la capa delgada de la Tierra donde se desarrolla la vida, abarcando la…
La botánica es la disciplina estudia las plantas. Se ocupa del reino vegetal en un sentido amplio,…
La célula es la unidad estructural, funcional y biológica más pequeña de todos los seres vivos. Es considerada…
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
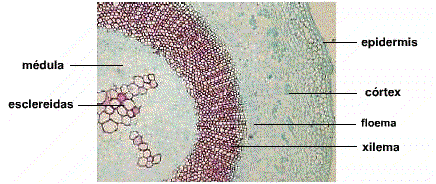{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}