
Los insectos
Los insectos son animales invertebrados, (sin huesos) que pertenecen al grupo de artrópodos son los organismos más numerosos y diversos del mundo animal.
La palabra “insecto del latín insectum, significa” animal dividido en secciones. En efecto, la clase de los insectos forma parte de la serie de los artrópodos, que son animales de cuerpo segmentado. El cuerpo del insecto comprende tres partes:
- La cabeza
- El tórax
- El abdomen
Todos ellos protegidos por un esqueleto externo de quitina.
Exoesqueleto
El exoesqueleto o ectoesqueleto es el esqueleto externo que recubre todo el cuerpo de los insectos y demás artrópodos y que también se conoce como integumento. En insectos está formado por una sucesión de capas; de adentro hacia afuera estas son: la membrana basal, la epidermis o hipodermis y la cutícula; la única capa celular es la epidermis; el resto no posee células y está compuesto por algunas de las siguientes sustancias: quitina, artropodina, esclerotina, cera y melanina. El componente rígido, la esclerotina, cumple varios papeles funcionales que incluyen la protección mecánica del insecto y el apoyo de los músculos esqueléticos, a través del llamado endoesqueleto; en los insectos terrestres, el exoesqueleto también actúa como una barrera para evitar la desecación o pérdida del agua interna. El exoesqueleto apareció por primera vez en el registro fósil hace unos 550 millones de años y su evolución ha sido crítica para la radiación adaptativa y la conquista de casi todos los nichos ecológicos del planeta que los artrópodos han venido realizando desde el Cámbrico
La cabeza
Está formada por cuatro segmentos íntimamente soldados entre sí, lleva un par de ojos compuestos o facetados, acompañado de ojos simples u ocelos en número variable: un par de antenas y los órganos bucales adaptados a la presión y a la masticación

La cabeza es la región anterior del cuerpo, en forma de cápsula, que contiene los ojos, antenas y piezas bucales. La forma de la cabeza varía considerablemente entre los insectos para dar espacio a los órganos sensoriales y a las piezas bucales. La parte externa endurecida o esclerosada de la cabeza se llama cráneo.
La cabeza de los insectos está subdividida por suturas en un número de escleritos más o menos diferenciados que varían entre los diferentes grupos. Típicamente hay una sutura en forma de «Y» invertida, extendiéndose a lo largo de la parte dorsal y anterior de la cabeza, bifurcándose por encima del ocelo para formar dos suturas divergentes, las cuales se extienden hacia abajo en los lados anteriores de la cabeza. La parte dorsal de esta sutura (la base de la Y) es llamada sutura coronal y las dos ramas anteriores suturas frontales. Por otra parte, la cabeza de los insectos está constituida de una región preoral y de una región postoral. La región preoral contiene los ojos compuestos, ocelos, antenas y áreas faciales, incluido el labio superior, y la parte postoral contiene las mandíbulas, las maxilas y los labios.
Internamente, el exoesqueleto de la cápsula cefálica de los insectos se invagina para formar las ramas del tentorio que sirven como sitios de inserción muscular.
Ojos

Ojos compuestos de un odonato.
La mayoría de los insectos tienen un par de ojos compuestos relativamente grandes, localizados dorso-lateralmente en la cabeza. La superficie de cada ojo compuesto está dividida en un cierto número de áreas circulares o hexagonales llamadas facetas u omatidios; cada faceta es una lente de una única unidad visual. En adición a los ojos compuestos, la mayoría de los insectos posee tres ojos simples u ocelos localizados en la parte superior de la cabeza, entre los ojos compuestos

Anatomía de un insecto
- Antena.
- (inferior)
- Ocelos (superior)
- Ojos compuestos
- Ganglio cerebral (Sistema nervioso (insectos)
- Protórax
- Vaso sanguíneo dorsal
- Tráquea. (Tronco con espiráculo)
- Mesotórax
- Metatórax
- Ala anterior
- Ala posterior
- Intestino medio (mesenterón)
- Tubo dorsal (corazón)
- Ovario
- Intestino inferior (proctodeo) (intestino grueso, recto y ano)
- Ano
- Oviducto
- Terminaciones nerviosas (ganglios abdominales)
- Túbulos de Malpighi
- Almohadilla del tarso
- Garras
- Fémur.
- Trocánter.
- Intestino superior (estomodeo)
- Ganglio Torácico
- Glándula Salivar
- Ganglio subesofágico
- Piezas bucales
Antenas
La antena funciona como un correo casi exclusivamente en la percepción sensorial, algunos de los usos sensoriales de antenas incluyen el movimiento y la orientación, olor, sonido, humedad, y una variedad de señales químicas. Las antenas varían mucho entre los insectos, pero todas siguen un plan básico, el primer y segundo segmentos de los cuales se denomina el escapo y pedicelo, respectivamente. Los segmentos o flagelómeros antenales restantes se denominan conjuntamente el flagelo Hay muchos tipos de antenas en las diferentes especies, incluidos seta, moniliforme, aserrada, pectinada, Clavate, laminada, Geniculada y plumosa los

- ‘Aristada’ Las antenas son de bolsa, con una cerda lateral. (Por ejemplo, Díptera)
- ‘Filiforme’ Las antenas son comunes entre muchos géneros diferentes, que tienen una forma simple, en forma de hilo.
- ‘Cerdosa’. Las antenas tienen muchas articulaciones. La antena se estrecha gradualmente desde la base hasta la punta (por ejemplo, Thysanura, Blattodea, Efemerópteros, Plecoptera y Trichoptera).
- ‘Moniliforme’. Las antenas tienen segmentos en forma de un collar de cuentas (por ejemplo, Coleóptera)
- ‘Aserrada’. Las antenas tienen segmentos que están en ángulo en un lado, dando la apariencia de un borde de sierra (por ejemplo, Coleóptera).
- ‘Pectinada’. Las antenas tienen segmentos que son más largos en un lado, dando la apariencia de un peine (por ejemplo Hemynoptera -. Symphyta y Coleóptera).
- ‘Clavate’. Las antenas tienen segmentos que se vuelven más amplios hacia la punta. Este puede ser gradual a lo largo de su longitud, o un aumento repentino, por lo que afecta principalmente a los últimos articulaciones, dando la apariencia de un garrote (por ejemplo, lepidópteros y Coleóptera),
- ‘Laminar’. Las antenas tienen segmentos que hacia el final son aplanados y la placa da la apariencia de un ventilador hacia el final de las antenas (por ejemplo, Coleóptera – Scarabaeidae.).
- ‘Geniculada’. Las antenas tienen una brusca curva o codo a lo largo de la antena (por ejemplo, Hymenoptera -. Formicidae y Coleóptera).
- ‘Plumosa’ cada segmento de la antena tiene un número de ramas finas, filiformes. Esto da la apariencia de una pluma (por ejemplo, Díptera)
Piezas Bucales
Las piezas bucales de insectos consisten en el maxilar superior, labio, y en algunas especies, las mandíbulas El labrum es un sencillo, esclerito fundido, a menudo llamado el labio superior, y se mueve longitudinalmente, que está articulada a la clípeo. Las mandíbulas (quijadas) son un par altamente esclerotizado de las estructuras que se mueven en ángulo recto con el cuerpo, que se utiliza para morder, masticar, y cortar alimentos. Los maxilares son estructuras pareadas, que también se pueden mover en ángulo recto con el cuerpo y poseer palpos segmentados. El labio (labio inferior) es la estructura fundida que se mueve longitudinalmente y posee un par de palpos segmentados.

Evolución de las piezas bucales de insectos a partir de
- las más primitivas, masticadoras de un saltamontes, en el centro), a
- tipo lamedor de una abeja,
- tipo de sifón de una mariposa, y
- tipo de succión de un mosquito hembra.
Leyenda: a – antenas
c – ojo compuesto
lb – labium
lr – labrum
md – mandíbulas
mx – maxilas
Las piezas bucales pueden tener por lo menos tres posiciones diferentes según la posición de la cabeza: prognatas, opistognatas, o hipognatas. En especies con prognatismo, la cabeza se coloca verticalmente alineada con el cuerpo, tales como especies de Formicidae; mientras que en las hipognatas, la cabeza está alineada horizontalmente adyacente al cuerpo. Una cabeza opistognata se coloca en diagonal, como las especies de Blattodea y algunos Coleóptera Las piezas bucales varían mucho en diferentes órdenes de insectos, pero los dos grupos funcionales principales son mandibuladas y haustellate. Piezas bucales haustellate se utilizan para la aspiración de líquidos y pueden ser clasificados por la presencia de estiletes, que incluyen la perforación, chupar, esponja, y sifón. Los estiletes son proyecciones en forma de aguja que se utilizan para penetrar los tejidos de plantas y animales. Los estiletes y la sonda de alimentación forman las mandíbulas modificadas, el maxilar y la hipofaringe.
- Mandibulados Tienen piezas bucales, entre los más comunes en los insectos, se utilizan para morder y moler los alimentos sólidos.
- Chupadores bucales tienen estiletes, y se utilizan para penetrar en el tejido sólido y después aspirar alimentos líquidos.
- Esponja Las piezas bucales se utilizan para la esponja y chupar los líquidos, y estiletes falta (por ejemplo, la mayoría Díptera).
- Sifón Los aparatos bucales carecen de estiletes y se utilizan para succionar los líquidos, y se encuentran comúnmente entre las especies de Lepidóptera.
Las piezas bucales mandibulares se encuentran en especies de Odonata, Isóptera, adulto Neuróptera,
Coleóptera, Hymenoptera, Blattaria, Orthoptera, y Lepidóptera. Sin embargo, la mayoría de adultos de lepidópteros tienen piezas bucales, mientras que sus larvas (comúnmente llamadas orugas ) tienen mandíbulas.
Mandíbulas
El labro es un amplio lóbulo que forma el techo de la cavidad preoral, suspendido del clípeo en frente de la boca y que forma el labio superior. En su lado interior, es membranosa y puede ser producido en un lóbulo medio, el epifaringe, teniendo algunos sensilias. El labrum se levanta lejos de las mandíbulas por dos músculos que surgen en la cabeza y se inserta medialmente en el margen anterior del labrum. Está cerrado contra las mandíbulas en parte por dos músculos que surgen en la cabeza y se inserta en los márgenes laterales posteriores en dos pequeños escleritos, la tormae, y, al menos en algunos insectos, por un resorte de resilina en la cutícula en la unión de la labrum con el clípeo. Uso diferencial de los músculos. Hasta hace poco, el labrum en general se considera asociada con el primer segmento de la cabeza. Sin embargo, estudios recientes de la embriología, la expresión génica, y la inervación del labrum muestran que está inervado por el tritocerebrum del cerebro, que son ganglios fusionados del tercer segmento de la cabeza. Este se forma a partir de la fusión de partes de un par de apéndices ancestrales que se encuentra en el segmento de tercera cabeza, mostrando su relación. Su ventral, o interior, de la superficie es generalmente membranosa y forma el lóbulo-. Como epifaringe, que lleva mechanosensilla y chemosensilla

Los insectos masticadores tienen dos mandíbulas, una a cada lado de la cabeza. Las mandíbulas están colocadas entre el labro y las maxilas. Las mandíbulas cortan y trituran los alimentos, y pueden ser utilizados para la defensa; en general, tienen un borde de corte apical, y la zona de los molares más basales muele la comida. Pueden ser extremadamente duras (alrededor de 3 en Mohs, o una dureza de penetración de alrededor de 30 kg / mm 2 ). Por lo tanto, muchas termitas y escarabajos no tienen ninguna dificultad física en perforar a través de láminas fabricadas con metales comunes como el cobre, el plomo, el estaño y el zinc. Suelen reforzarse mediante la adición de zinc, manganeso, o rara vez, el hierro, en cantidades de hasta un 4% del peso seco. Por lo general son las piezas bucales más grandes de los insectos masticadores, que se utilizan para masticar (corte, desgarro, aplaste, mastique) productos alimenticios. Se abren hacia el exterior (a los lados de la cabeza) y se unen medialmente. En insectos masticadores carnívoros, las mandíbulas pueden ser modificadas para ser más similar a un cuchillo, mientras que en los insectos masticadores herbívoros son más típicamente ancho y plano en sus caras opuestas (por ejemplo, orugas). En los escarabajos ciervo machos, las mandíbulas se modifican en un grado tal que no sirven ninguna función de alimentación, sino que se utilizan para defender los sitios de apareamiento de otros machos. En hormigas, las mandíbulas también cumplen una función defensiva (en particular en las castas de soldado). En las hormigas toro, las mandíbulas son largas y dentada, utilizadas como apéndices para la caza y defensa.
Situadas debajo de las mandíbulas, a la par maxilar e manipular alimentos durante la masticación. Los maxilares pueden tener pelos y «dientes» a lo largo de sus márgenes interiores. Al margen exterior, la gálea es una forma de copa o de la estructura de la cucharada-como, que se sienta sobre el borde exterior del labio. También tienen palpadores, que se utilizan para detectar las características de los alimentos potenciales. Los maxilares ocupan una posición lateral, uno a cada lado de la cabeza detrás de las mandíbulas. La parte proximal del maxilar superior consta de un cardo basal, que tiene una única articulación con la cabeza, y una placa plana, las estipes, articuladas al cardo. Tanto el cardo como las estipes están débilmente unidas a la cabeza por la membrana por lo que son capaces de movimiento. Distal sobre los estipes son dos lóbulos, un lacinia interior y una exterior Gálea, uno o ambos de los cuales puede estar ausente. Más lateralmente en los estipes hay un compuesto articulado por un número de segmentos; en ortópteros hay cinco que se insertan en el cardo, y los músculos aductores ventrales surgidas en la tienda del cerebelo se insertan tanto en cardo y estípites. Con origen en los estípites son los músculos flexores de lacinia y gálea y otro flexor lacineal surge en el cráneo, pero ni la lacinia ni la gálea tienen un músculo extensor. El palpo tiene elevador del ano y músculos depresores que se plantean en los estípites, y cada segmento del palpo tiene un solo músculo que causa la flexión del siguiente segmento.
En partes de la boca, el labio es una estructura cuadrúpeda, aunque se forma a partir de dos anillos condensados maxilares secundaria. Esto puede ser descrito como el piso de la boca. Con los maxilares, ayuda con la manipulación de los alimentos durante la masticación o, en el caso inusual de la libélula ninfa que se extiende hacia fuera para jalas la presa de nuevo a la cabeza, donde las mandíbulas pueden comerlo. El labio es similar a la estructura del maxilar, pero con los apéndices de los dos lados fusionados por la línea media, por lo que vienen a formar una placa de mediana. La parte basal del labio, equivalente a los Cardines maxilares y posiblemente incluyendo una parte del esternón del segmento labial, se llama la postmentum. Esto puede ser subdividido en un submentum proximal y un mentón distal. Distal a la postmentum, y equivalente a los estípites maxilares fusionados, es el prementum. El prementum cierra la cavidad preoral desde atrás. Terminal, lleva cuatro lóbulos, dos glosas interiores, y dos paraglossae exterior, que se conocen colectivamente como el lígula. Uno o ambos pares de lóbulos pueden estar ausentes o pueden estar fusionados.
La hipofaringe es un lóbulo medio inmediatamente detrás de la boca, que se proyecta hacia delante desde la parte posterior de la cavidad preoral; se trata de un lóbulo de origen incierto, pero tal vez asociado con el segmento mandibular; en apterygotes, tijeretas, y las efímeras ninfales, la hipofaringe lleva un par de lóbulos laterales, el superlinguae (singular: superlingua ). Se divide la cavidad en una bolsa de alimentos dorsal, o cibarium, y una salivarium ventral en la que se abre el conducto salival. Se encuentra comúnmente fusionado al libium. La mayor parte de la hipofaringe es membranosa, pero la cara adoral se esclerotizados distal y proximal contiene un par de escleritos suspensorios que se extienden hacia arriba para terminar en la pared lateral de la estomodeo. Los músculos que surgen en el frons se insertan en estos escleritos, que distalmente están articuladas a un par de escleritos linguales. Estos, a su vez, se han insertado en los pares de los músculos que se producen al tentorium y labio. Los diversos músculos sirven para balancear los delanteros hipofaringe y la espalda, y en la cucaracha, dos músculos más corren a través de la hipofaringe y dilatan el orificio salival y ampliar la salivarium.
Ejemplo de mandíbulas en insectos sifón




Sifón
La probóscide o probosces (plural – probóscides) se forman a partir de maxilar galeae y es la adaptación se encuentra en algunos insectos para chupar Los músculos de la faringe o cibarium están fuertemente desarrollados y constituyen la bomba. En Hemíptera y muchos Díptera, que se alimentan de los fluidos dentro de las plantas o los animales, algunos componentes de las piezas bucales se modifican para la perforación, y las estructuras alargadas son llamadas estiletes. Las estructuras tubulares combinados se conocen como la proboscis, aunque la terminología especializada se utiliza en algunos grupos.
En especies de lepidópteros, que consta de dos tubos unidos por ganchos y separable para la limpieza. Cada tubo es cóncava hacia dentro, formando así un tubo central a través del cual se aspira la humedad. La aspiración se realiza a través de la contracción. Ellos brillan y cinco segmentados en algunas de las familias más basales, y con frecuencia se doblan. El. forma y dimensiones de la probóscide han evolucionado para dar diferentes especies de más amplia y por lo tanto, las dietas más ventajosas Hay una alométrico relación de escala entre la masa corporal de los lepidópteros y la longitud de la probóscide de la que una salida de adaptación interesante es la inusualmente larga de lengua polilla halcón Xanthopan morgani praedicta. Charles Darwin predicho la existencia y la probóscide longitud de esta polilla antes de su descubrimiento basado en su conocimiento de la tan estimulado orquídea estrella de Madagascar Angraecum sesquipedale
Esponjas
Las piezas bucales de los insectos que se alimentan de los fluidos son modificados de diversas maneras para formar un tubo a través del cual el líquido pueda mezclarse en la boca y por lo general otra a través del cual pasa la saliva. Los músculos de la faringe o cibarium se desarrollan fuertemente para formar una bomba. En moscas no-mordedoras, las mandíbulas están ausentes y otras estructuras se reducen.; los palpos labiales se han modificado para formar el labelo y los palpos maxilares están presentes, aunque a veces corto. En Brachycera, el labelo es especialmente destacado y utilizarse para la esponja alimento líquido o semilíquido. El labelo es una estructura compleja que consta de muchos surcos, llamado pseudotrachea, que son líquidos o secreciones salivales de la labella ayudar a disolver y recoger las partículas de alimentos para que puedan ser más fácilmente absorbidos por el pseudotracheae; esto se cree que se producen por la acción capilar. El alimento líquido se extrae a continuación, a partir de la pseudotracheae a través del canal de alimentación en el esófago. Las piezas bucales de las abejas son de masticación y de recubrimiento para la succión. El pulido es un modo de alimentación en la que el líquido o semilíquido adherente de alimentos a un órgano protrusible, o «lengua», se transfiere de sustrato a la boca. En el abeja de la miel (Hymenoptera: Apidae: Apis mellifera), forman el glosas labial alargada y fusionado una lengua vellosa, que está rodeado por el maxilar galeae y los palpos labiales para formar una trompa tubular que contiene un canal de alimentación. En la alimentación, la lengua se sumerge en el néctar o miel, que se adhiere a los pelos, y luego se retrae de manera que el líquido adherido se realiza en el espacio entre la galeae y palpos labiales. Este movimiento glossal de ida y vuelta se produce repetidamente. Movimiento de líquido a la boca resulta aparentemente de la acción de la bomba cibarial, facilitado por cada uno de retracción de la lengua empujando líquido hasta el canal de alimentos.
Protórax
El protórax está protegido por un gran pronoto que se extiende desde el cuello en el lado anterior para cubrir el mesotórax y la mayor parte del metatórax en el lado posterior, así como cubrir la pleura en el protórax. El pronoto del protórax puede ser simple en estructura y pequeño en comparación con la otra nota, pero en los escarabajos, mantis, muchos errores, y algunos ortópteros, pronoto está expandida, y en las cucarachas, se forma un escudo que cubre parte de la cabeza y el mesotórax.
Pterotórax
Debido a que el mesotórax y metatórax sostienen las alas, tienen un nombre combinado llamado protórax (pteron=ala). El ala delantera, que se conoce con diferentes nombres en diferentes órdenes (por ejemplo, la tegminas en ortópteros y élitros en Coleóptera), surge entre el mesonoto y el mesopleura, y las alas posteriores se articulan entre el metanoto y la metapleura. El segundo y tercer par de patas emergen de la mesopleura y metapleura. El mesotórax y el metatórax cada uno tiene una sutura pleural (suturas mesopleural y metapleural) que se extienden desde la base del ala a la coxa de la pata. El esclerito anterior a la sutura pleural se llama la episternum (en orden, mesepisternum y metepisternum). El esclerito posterior a la sutura se llama la epimiron (en orden, mesepimiron y metepimiron). Espiráculos, los órganos externos del sistema respiratorio, se encuentran en el Pterotórax, por lo general uno entre el pro y mesopleuron, así como uno entre el meso y metapleuron.
La región ventral o esternón sigue la misma composición, con el proesterno bajo el protórax, meso bajo el mesotórax y metasterno bajo el metatórax. El notum, la pleura y el esternón de cada segmento tienen una variedad de diferentes escleritos y suturas, que varían en diferentes órdenes, y que no serán discutidos en detalle en esta sección.
Ala (insectos)

La característica filogenéticamente más avanzada de los insectos son los dos pares de alas situadas en el segundo y tercer segmentos torácicos. Los insectos son los únicos invertebrados que han desarrollado tal capacidad, y esto ha jugado un papel importante en su éxito. El vuelo del insecto no se ha comprendido muy bien debido a la dependencia de los efectos aerodinámicos turbulentos. Los grupos de insectos primitivos utilizan los músculos que actúan directamente sobre la estructura del ala. Los grupos más avanzados que integran los Neoptera tienen alas plegables, y sus músculos que actúan en la pared del tórax y el poder de las alas indirectamente. Estos músculos son capaces de contraerse varias veces por cada impulso nervioso, permitiendo que las alas batan más rápido que lo que sería normalmente posible.
El vuelo del insecto puede ser extremadamente rápido, maniobrable y versátil, posiblemente debido a la forma cambiante, control extraordinario, y el movimiento variable del ala del insecto. Diferentes órdenes de insectos utilizan diferentes mecanismos de vuelo; por ejemplo, el vuelo de una mariposa se puede explicar mediante el estado de equilibrio y fina teoría aerodinámica.
Anatomía interna

Cada una de las alas consiste en una membrana delgada soportada por un sistema de venas. La membrana está formada por dos capas estrechamente yuxtapuestas, mientras que las venas se forman donde las dos capas permanecen separadas y las cutículas pueden ser más gruesas y más fuertemente esclerotizadas. Dentro de cada una de las venas principales hay un nervio y una tráquea, y, puesto que las cavidades de las venas están conectados con el hemocele, la hemolinfa puede fluir hacia las alas. También, el lumen del ala, siendo una extensión del hemocele, contiene las tráqueas, los nervios y la hemolinfa. A medida que se desarrolla el ala, el dorsal y capas integumental ventrales se convierten estrechamente apposed sobre la mayor parte de su área, formando la membrana del ala. Las áreas restantes forman canales, las venas, los futuros en que puedan formarse los nervios y tráqueas. La cutícula que rodea las venas se vuelve más gruesa y más fuertemente esclerotizado para proporcionar resistencia y rigidez a la banda. Pelos de dos tipos pueden ocurrir en las alas: microtriquios, que son pequeñas y dispersas irregularmente, y macrotrichia, que son más grandes, socketed, y puede ser restringida a las venas. Las escalas de Lepidoptera y Trichoptera son altamente macrotrichia modificados.
Venas

Venación de insectos alados, basado en el sistema de Comstock-Needham
En algunos insectos muy pequeños, la nervadura puede reducirse en gran medida. En avispas chalcid, por ejemplo, solo la subcosta y parte del radio están presentes. A la inversa, un aumento de la nervadura se puede producir por la ramificación de venas existentes para producir venas accesorias o por el desarrollo de, venas intercalares adicionales entre los originales, como en las alas de Orthoptera (saltamontes y grillos). Un gran número de venas transversales están presentes en algunos insectos, y pueden formar un retículo como en las alas de los Odonata(libélulas y caballitos del diablo) y en la base de las alas anteriores de Tettigonioidea y Acridoidea (saltamontes y langostas, respectivamente).
El archedictyon es el nombre dado a un esquema hipotético de venación alar propuesto para el primer insecto alado. Se basa en una combinación de la especulación y los datos fósiles. Dado que se cree que todos los insectos alados han evolucionado de un ancestro común, el archediction representa la «plantilla» que ha sido modificado (y aerodinámica) por selección natural durante 200 millones de años. Según el dogma actual, el archedictyon contenía seis a ocho venas longitudinales. Estas venas (y sus ramas) se denominan de acuerdo con un sistema ideado por John y George Comstock-Needham el sistema de Comstock-Needham:
- Costa (C) – Borde del ala principal
- Subcosta (Sc) – Segunda vena longitudinal (detrás de la costa costa), no ramificada
- Radio (R) – Tercera vena longitudinal, de una a cinco ramas llegan a ser el margen del ala
- Media (M) -Cuarta vena longitudinal, de una a cuatro ramas están cerca del margen del al
- Cúbito (Cu) – Quinta vena longitudinal, de una a tres ramas son parte del margen del ala
- Venas anales (A1, A2, A3) – Venas no ramificadas detrás del cúbito
La costa (C) es la vena marginal líder en la mayoría de los insectos, a pesar de una pequeña vena, la precosta, a veces se encuentra por encima de la costa. En casi todos los insectos existentes, la precosta se funde con la costa; la costa casi nunca ramas porque está en el borde de ataque, que se asocia en su base con la placa del húmero. La tráquea de la vena costal es quizás una rama de la tráquea subcostal. Está situado detrás de la costa es la tercera línea, el subcosta, que se ramifica en dos venas separado: el anterior y posterior. La base de la subcosta se asocia con el extremo distal del cuello de la primera axilar. La cuarta vena es el radio, que está ramificado en cinco venas separadas. El radio es generalmente más fuerte de la vena del ala. Hacia la mitad de la banda, se bifurca en una primera rama indivisa (R1) y una segunda rama, llamada el sector radial (Ra), que subdivide dicotómica en cuatro ramas distales (R2, R3, R4, R5). Basales, el radio se forma flexible unida con el extremo anterior de la segunda axilar (2Ax)
La quinta vena del ala es el medio de comunicación. En el patrón arquetipo (A), las horquillas de los medios de comunicación están divididas en dos ramas principales, una anterior a los medios de comunicación (MA), que se dividen en dos ramas distales (MA1, MA2), y un sector de la mediana, o posterior de los medios de comunicación (MP), que tiene cuatro ramas terminales (M1, M2, M3, M4). En la mayoría de los insectos modernos, la parte anterior de medios se ha perdido, y los «medios» usualmente son parte posterior media de cuatro ramas con el tallo basal común. En el Ephemerida, de acuerdo con las interpretaciones actuales de la venación alar, ambas ramas de los medios de comunicación se conservan, mientras que en los odonatos, los medios de comunicación persisten en la rama anterior primitiva. El vástago de los medios de comunicación a menudo se une con el radio, pero cuando se produce como una vena distinta, su base está asociada con la placa de mediano distal que está esclerotizada continuamente con este último. El cúbito, la sexta vena del ala, es principalmente de dos ramificaciones. La bifurcación primaria se lleva a cabo cerca de la base del ala, formando las dos ramas principales (CU1, Cu2). La rama anterior puede dividirse en varias ramas secundarias, pero comúnmente se bifurca en dos ramas distales. La segunda rama del cúbito (Cu2) en Hymenoptera, Trichoptera y Lepidoptera, fue confundida por Comstock y Needham como la primera vena anal. El proximal es el tallo principal del cúbito está asociada con la placa mediano distal (m ‘) de la base del ala.

El Postcubito (Pcu) es el primer sistema anal de Comstock y Needham. El Postcúbito, sin embargo, ha tenido el estatus de ala con vena independiente y debería ser reconocida como tal. En ala de ninfas, la tráquea sube entre el traque cubital y un grupo de venas en la tráquea. En las alas maduras de insectos más generales, el postcúbito está siempre asociado por su proximidad con el cubito, y nunca está conectado al flexor esclerital (3Ax) de la base del ala. En Neuróptera, Mecoptera, y Trichoptera, el postcúbito puede estar más estrechamente asociado a un conjunto de venas, pero su base está siempre libre posteriormente. El postcúbito no está usualmente en ramificaciones; primeramente, está formado por dos ramificaciones. El conjunto de venas (lV a nV) son las venas anales inmediatamente asociadas con el tercer axilar, el cual está directamente afectado al movimiento de su esclerito y que están directamente afectados por el movimiento de este esclerito que provoca la flexión de las alas. En número, las venas varían de una a 12, de acuerdo con la ampliación de la zona del ala. Las tráqueas suelen surgir de una raíz común traqueal en los insectos ninfas y las venas son consideradas como ramas de una única vena anal. Distalmente, las venas son ya sea simples o ramificadas. La vena yugal (J) del lóbulo yugal del ala es a menudo ocupada por una red de venas irregulares, o puede ser completamente membranosa; a veces contiene uno o dos distintas, pequeñas venas, la primera vena yugal, o de la vena arcuata, y la segunda vena yugal, o de la vena cardinales(2J).

- C-Sc Venas cruzadas – Corre entre la costa y la subcosta
- R Venas cruzadas – Corre entra la raíces adyacentes y el radio
- R-M Venas cruzadas – Corre entre el radio y la media
- M-Cu Venas cruzadas – Corre entre la media y el cubito
Todas las venas del ala están sujetas a la bifurcación secundaria de la unión de venas transversales. En algunos órdenes de insectos, el cruce de las venas es tan numerosas que todo el patrón se convierte en una estrecha red de venas ramificadas y venas transversales. Normalmente, un número definido de venas cruzadas tienen lugares específicos se producen. Las más constantes venas cursadas son el húmero cruzada vena (h) entre la costa y subcosta, el radial transversal de la vena (r) entre R y el primer tenedor de rupias, la sectorial transversal de la vena (s) entre las dos horquillas de R8, la mediana de los vena (mm) entre M2 y M3, y la cruz de la vena (m-cu) mediocubital entre los medios de comunicación and the cubitus. Las venas de las alas de los insectos se caracterizan por una colocación convexo-cóncava, tales como los que se observan en las efímeras (es decir, cóncava es «hacia abajo» y convexa es «arriba») por su ramificación alternativa. Cada vez que hay una vena en una horquilla siempre hay un interpolado de la posición opuesta entre las dos ramas. La vena cóncava se bifurcará en dos venas cóncavas (con la vena interpolada convexo) y la alteración periódica de la venas está preservado Las venas del ala parecen caer en un patrón ondulado en función de si tienen una tendencia a plegarse hacia arriba o hacia abajo cuando se relaja el ala. Los ejes basales de las venas son convexas, pero cada uno horquillas vena distalmente en una rama convexa anterior y una rama cóncava posterior. Por lo tanto, la Costa y subcostal se consideran como convexas y ramas cóncavas de una primera vena primaria, Rs es la rama cóncava del radio, los medios de comunicación posteriores la rama cóncava de los medios de comunicación, Cu1 y Cu2, son respectivamente convexas y cóncavas, mientras que el postcúbito primitiva y el primer grupo de venas tienen cada una rama anterior convexa y una rama cóncava posterior. La naturaleza convexa o cóncava de las venas se ha utilizado como evidencia en la determinación de la identidad de la persistencia de las ramas distales de las venas de los insectos modernos, pero no se ha demostrado que ser coherente para todas las alas.
Campos
El área de las alas está delimitado y se subdividen por pliegues, a lo largo de los cuales las alas pueden plegarse, y tener líneas de flexión para el vuelo. Entre la flexión y las líneas de plegado, la distinción fundamental es a menudo borrosa, como las líneas de pliegue pueden permitir cierta flexibilidad o viceversa. Dos constantes, que se encuentran en casi todas las alas de los insectos, son el Claval (una línea de flexión) y yugal pliegues (o línea de doblez), formando límites variables y poco satisfactorios. Plegamientos ala puede muy complicado, con plegado transversal que ocurre en las alas posteriores de Dermáptera y Coleóptera, y en algunos insectos, la zona anal puede plegarse como un abanico. Los cuatro campos diferentes que se encuentran en las alas de los insectos son:
- Remigio
- Área anal
- Área jugal
- Área axilar
- Álula
El vano está bordeado por el pliegue vanal, que normalmente se produce entre el postcúbito y la primera vena vanal. Los ortópteros, por lo general tienen esta posición. En las alas anteriores de los Blattidae, sin embargo, el único pliegue en esta parte del ala se encuentra inmediatamente antes de la postcúbito. En los plecóptera, el pliegue vanal es posterior al postcubital, pero próximamente se cruza la base de la primera vena vanal. En la cigarra, el pliegue vanal se encuentra inmediatamente detrás de la primera vena vanal (lV). Estas pequeñas variaciones en la posición real del pliegue vanal, sin embargo, no afectan a la unidad de acción de las venas vanales, controladas por el flexor esclertial (3AX), en la flexión del ala. En las alas posteriores de la mayoría de los ortópteros, unos venas secundarias forman una costilla en el pliegue vanal. El vanos generalmente se forma triangularmente con sus venas típicamente hacia fuera desde el tercer axilar como las varillas de un abanico. Algunas de las venas vanal pueden estar ramificadas, y las venas secundarias pueden alternar con las venas principales. La región vanal generalmente se desarrolla mejor en las alas posteriores, en la que puede ser ampliada para formar una superficie sustentadora, como en Plecoptera y Orthoptera. Las grandes expansiones en forma de abanico de las alas posteriores de Acrididae son claramente las regiones vanalws, ya que sus venas están soportados en los terceros escleritos axilares en la base de las alas, aunque Martínov (1925) atribuye la mayor parte de las áreas de fans en Acrididae a las regiones yúgales de las alas. El verdadero jugum del ala está representada solo por la pequeña membrana (Ju) de la última vena vanal. El jugum está más desarrollado en alguna otra ortópteros, como en el Mantidae. En la mayoría de los insectos más altas con alas estrechas, la venal vanal se reduce, y el pliegue vanal se pierde, pero incluso en estos casos, el ala de flexión puede doblarse a lo largo de una línea entre el postcúbito y la primera vena vanal.
La región yugal o neala, es una región del ala que es por lo general una pequeña zona membranosa proximal a la base del vannus reforzada por unos engrosamientos-vena como pequeñas e irregulares; pero cuando bien desarrollada, es una sección distinta de la banda y puede contener uno o dos venas yugal. Cuando el área malar del ala anterior se desarrolla como un lóbulo libre, se proyecta por debajo del ángulo humeral de las alas posteriores y por lo tanto sirve para uncir las dos alas juntas. En el grupo Jugatae de Lepidoptera, lleva un lóbulo de dedo largo. La región yugal se denominó neala («a la nueva»), ya que es evidente que una parte secundaria y recientemente desarrollado del ala. La región axilar que contiene los Escleritos axilares tiene, en general, la forma de un triángulo escaleno. La base del triángulo (ab) es la bisagra del ala con el cuerpo; el vértice (c) es el extremo distal de la tercera esclerito axilar; el lado más largo es anterior hasta el ápice. El punto D en el lado anterior del triángulo marca la articulación de la vena radial con el segundo esclerito axilar. La línea entre d y c es la basal plica (bf) o pliegue del ala en la base del campo mediocubital. En el ángulo posterior de la base del ala en algunos dípteros hay un par de lóbulos membranosos (escamas, o calypteres) conocidos como la álula. El álula está bien desarrollada en la mosca doméstica. La escama del exterior (c) surge de la base del ala detrás de la tercera sclerite axilar (3AX) y, evidentemente, representa el lóbulo yugal de otros insectos (A, D); la escama interno más grande (d) surge de la margen posterior escutelar del tergo del segmento del ala de soporte y forma una cubierta protectora, el capó como el cabestro. En el ala flexión, la escama exterior del álula se pone de cabeza por encima de la escama del interior, no siendo este último afectado por el movimiento del ala. En muchos Díptera, una profunda incisión en el área anal de la membrana del ala detrás de la única vena vannal pone en marcha una alar proximal distal del lóbulo de la escama exterior del álula.
Articulaciones

Los diversos movimientos de las alas, especialmente en los insectos que flexionan sus alas horizontalmente sobre la espalda cuando está en reposo, exigir una estructura articular más complicado en la base del ala que un mero bisagra del ala con el cuerpo. Cada ala está unida al cuerpo por un área basal membranosa, pero la membrana articular contiene un número de pequeñas Escleritos articulares, conocidos colectivamente como la pteralia. Los pteralia incluyen una placa humeral anterior en la base de la vena costal, un grupo de axilar (ax) asociado con el subcostal, radial, y venas vannal, y dos placas de menos definida mediana (M, M ‘) en la base del área mediocubital. Los axilares se desarrollan específicamente solo en los insectos ala-flexionar, donde constituyen el mecanismo flexor del ala operado por el músculo flexor de la que surge en la Pleurón. Características de la base del ala es también un pequeño lóbulo en el margen anterior de la zona articular proximal a la placa del húmero, que, en el ala anterior de algunos insectos, se desarrolla en una escala similar a la aleta grande, plana, la Tegula, la superposición la base del ala. Posteriormente, la membrana articular a menudo se forma un amplio lóbulo entre el ala y el cuerpo, y su margen es generalmente más gruesa y corrugado, dando la apariencia de un ligamento, el llamado cable de axilar, mesally continua con la parte posterior de plegado escutelar marginal de la placa de tergal que lleva el ala.
Los escleritos articulares o pteralia, de la base del ala de los insectos ala-flexionar y sus relaciones con el cuerpo y las venas de las alas, que se muestra de forma esquemática, son las siguientes:
- Placa humeral
- Axilar primera
- Axilar segunda
- Axilar tercera
- Axilar cuarta
- Humeral media (m, m’)
La placa del húmero suele ser un pequeño sclerito en el margen anterior de la base del ala, móvil y articulada con la base de la vena costal. Odonata han sus placas de húmero enlargened en gran medida, con dos músculos derivados de la episternum insertado en las placas de húmero y dos desde el borde de la epimeron insertado en la placa axilar. La primera sclerite axilar (LAX) es la placa de articulación anterior de la base del ala. Su parte anterior está soportado en el proceso de ala notal anterior del tergo (ANP); su parte posterior se articula con el margen de tergal. El extremo anterior de la esclerito se produce generalmente como un brazo delgado, el ápice de los cuales (e) siempre está asociado con la base de la vena subcostal (SC), a pesar de que no está unida con este último. El cuerpo de la esclerito articula lateralmente con el segundo axilar. El segundo sclerite axilar (2Ax) es más variable en la forma que la primera axilar, pero sus relaciones mecánicas no son menos definida. Se oblicuamente está articulada a la margen exterior del cuerpo de la primera axilar, y la vena radial (R) es siempre flexible unido a su extremo anterior (d). El segundo axilar presenta tanto un dorsal y un esclerotización ventral en la base del ala; su superficie ventral se basa en el proceso de ala del fulcro del Pleurón. El segundo axilar, por lo tanto, es el esclerito fundamental de la base del ala, y se manipula específicamente la vena radial.
La tercera sclerite axilar (3AX) se encuentra en la parte posterior de la región articular del ala. Su forma es muy variable y con frecuencia irregular, pero la tercera es la axilar sclerite en que se inserta el músculo flexor del ala (D). Mesally, se articula hacia delante (f) con el extremo posterior de la segunda axilar, y posteriormente (b) con el proceso de ala posterior de la tergo (PNP), o con un pequeño cuarto axilar cuando este último está presente. Distalmente, la tercera axilar se prolonga en un proceso siempre asociado con las bases del grupo de las venas en la región anal del ala, aquí denominado las venas vannal (V). La tercera axilar, por lo tanto, es por lo general la placa de bisagra posterior de la base del ala y es la esclerito activo del mecanismo de flexor, que manipula directamente las venas vannal. La contracción del músculo flexor (D) gira en la tercera axilar en sus articulaciones MESAL (b, f), y con ello levanta su brazo distal; este movimiento produce la flexión del ala. El cuarto sclerite axilar no es un elemento constante de la base del ala. Cuando está presente, por lo general es una placa pequeña que interviene entre la tercera axilar y el proceso de ala notal posterior, y es probablemente una pieza individual de este último.
Las placas de la mediana (m, m ‘) son también escleritos que no son tan definitivamente diferenciados como platos específicos como son los tres principales axilares, pero son elementos importantes del aparato flexor. Se encuentran en la zona mediana de la base distal del ala a las segunda y tercera axilar, y están separados entre sí por una línea oblicua (BF), que forma un prominente pliegue convexo durante la flexión del ala. La placa proximal (m) es por lo general unido al brazo distal de la tercera axilar y quizás debe considerarse como una parte de este último. La placa distal (m ‘) es menos constantemente presente como un esclerito distinta, y puede ser representado por un esclerotización general de la base del campo mediocubital del ala. Cuando las venas de esta región son distintos en sus bases, que están asociados con la placa de mediana exterior.
Acoplamiento, plegado, y otras características
En muchas especies de insectos, la proa y ala posterior se acoplan entre sí, lo que mejora la eficiencia aerodinámica de vuelo. El mecanismo más común de acoplamiento (por ejemplo, en Hymenoptera y Trichoptera) es una fila de pequeños ganchos en el margen delantero del ala posterior, o «hamuli», que bloquean en el ala delantera, manteniéndolos juntos (acoplamiento hamulate). En algunas otras especies de insectos (por ejemplo, los Mecoptera, Lepidoptera, y algunos Trichoptera) el lóbulo yugal del ala anterior cubre una parte del ala posterior (acoplamiento yugal), o de los márgenes de la proa y de la superposición trasera ala amplia (acoplamiento amplexiforme), o las cerdas alas posteriores, o frenillo, gancho bajo la estructura de contención o retinaluco en los anteriores. Cuando están en reposo, las alas caen sobre la parte posterior en la mayoría de los insectos, lo que puede implicar el plegado longitudinal de la membrana del ala y a veces también de plegado transversal. En algunas veces puede ocurrir a lo largo de las líneas de flexión. Aunque las líneas de plegado pueden ser transversales, como en las alas posteriores de escarabajos y tijeretas, que están normalmente radiales a la base del ala, permitiendo que las secciones adyacentes de un ala puedan ser plegadas encima o por debajo uno del otro. La línea de plegado más común es el pliegue malar, situada justo detrás de la tercera vena anal, aunque, la mayoría de los Neoptera tiene un espacio jugal justo detrás de la 3A vena de las alas anteriores. A veces también está presente en las alas posteriores. Cuando la zona anal de las alas posteriores es grande, como en un Orthoptera y Blattodea, la totalidad de esta parte puede ser plegada bajo la parte anterior de la banda a lo largo de un vanal y a veces un poco posterior al surco Claval. Además, en los Orthoptera y Blattodea, la zona anal se pliega como un abanico a lo largo de las venas, las venas anales pueden ser convexas, en las crestas de los pliegues, y la cóncava venas accesorias. Considerando que el surco Claval y pliegue malar son probablemente homólogas en diferentes especies, el pliegue vanal varía en su posición en diferentes taxones. Plegable es producida por un músculo que surge en el Pleurón y se inserta en el tercer esclerito axilar, cuando se contraen, los pivotes escleritales acercan sus puntos de articulación con el proceso notal posterior y el segunda esclerito axilar.
Como resultado, el brazo distal del tercer esclerito axilar gira hacia arriba y hacia el interior, de modo que, finalmente, su posición se invierte completamente. Las venas anales se articulan con este esclerito de tal manera que se mueven con él y se convierten en una parte flexionada el lomo del insecto. Esta actividad muscular en vuelo afecta a la potencia de salida del ala y por lo que también es importante en el control de vuelo. En los insectos orthopteroides, la elasticidad de la cutícula hace que el área vanal del ala se pliegue a lo largo de las venas. En consecuencia la energía se gasta en el despliegue de esta región cuando las alas se mueven a la posición de vuelo. En general, la extensión del ala probablemente se debe a la contracción de músculos vinculados a la esclerito basalar o, en algunos insectos, a la esclerito subalar.
La membrana que se encuentra entre las placas duras y las secciones del cuerpo en los artrópodos puede ser muy extensible, o plegada y laminada para proporcionar un menor grado de extensibilidad pero mayor grado de fuerza. De un estudio donde se examinaba el cuello de varias especies de libélulas para entender la forma y las funciones de la cutícula plegada, se extrajo que la membrana del cuello dispone de varios órdenes de pliegues. Este conjunto de estructuras permite a la propia cutícula a ser más rígida y tener mayores módulos elásticos porque los pliegues permiten que se deforme que si fuera lisa. Ya que se presenta a sólo en el lado externo, permite una flexibilidad mucho mayor en el sentido de la cara donde se encuentran los pliegues, ofreciendo una gran oposición al doblado en el sentido contrario.
Patas
Tipos de patas:
- Patas cursoriales modificadas para la carrera (escarabajo de tierra),
- Patas traseras saltadoras adaptadas para el salto (langosta),
- Patas delanteras fosoriales modificadas para cavar (grillo topo),
- Patas nadadoras modificadas para la natación (escarabajo del salto),
- Patas delanteras raptoriales modificadas para agarrar. Cazadores de presas (mantis),
- Patas de abejas obreras. (Abeja)
- Patas trepadoras / Prensiles / Con garra y modificadas para agarrar y trepar (piojos)

Partes de la pata:
- Coxa,
- Trocánter,
- Fémur,
- Tibia,
- Tarso
Los segmentos típicos y habituales de la pata de los insectos son la coxa, el trocánter, el fémur, la tibia, el pretarso y el tarso. La coxa en su forma más simétrica, tiene la forma de un corto cilindro o de cono truncado, aunque comúnmente es oval y puede ser casi esférica. El extremo proximal de la coxa está ceñida por una basicostal sutura submarginal que forma internamente una cresta, o basicosta, y pone en marcha una brida marginal, el coxomarginal o basicoxite. La basicosta fortalece la base de la coxa y se ensancha comúnmente en la pared exterior para dar inserción a los músculos; en el medio mesal de las coxas, sin embargo, por lo general es débil y a menudo confluentes con el margen coxal. Los músculos trocantéreo que tienen su origen en la coxa siempre están unidos distalmente a la basicosta. La coxa está unida al cuerpo por una membrana articular, el corium coxal, que rodea su base. Estas dos articulaciones son quizás los dorsales primarios y puntos articulares ventrales de la bisagra-subcoxo coxal. Además, la coxa del insecto tiene a menudo una articulación anterior con la anterior, extremo ventral de la trocantín, pero la articulación trochantinal no coexiste con una articulación esternal. La superficie articular pleural de la coxa es llevada en una inflexión mesal de la pared del coxal. Si la coxa es móvil sobre la articulación pleural sola, la superficie articular coxal está flexionada por lo general a una profundidad suficiente para dar una palanca para los músculos abductores insertados en el borde exterior de la base coxal. Distalmente la coxa lleva una articulación anterior y una posterior con el trocánter. La pared exterior de las coxas a menudo se caracteriza por una sutura que se extiende desde la base hasta la articulación trocánteres anteriores. En algunos insectos la sutura coxal está en consonancia con la sutura pleural, y en tales casos la coxa parece estar dividida en dos partes correspondientes a la episternum y epimeron de la Pleurón. La sutura coxal está ausente en muchos insectos.
La inflexión de la pared coxal teniendo la superficie articular pleural divide la pared lateral de la basicoxite en una parte prearticular y una parte postarticular, y las dos áreas a menudo aparecen como dos lóbulos marginales sobre la base de la coxa. El lóbulo posterior es por lo general el más grande y se denomina Meron.
El Meron puede ser ampliado en gran medida por una extensión distalmente en la pared posterior de la coxa; en el Neurópteros, Mecoptera, Trichoptera, y Lepidoptera, el Meron es tan grande que la coxa parece estar dividida en una pieza anterior, llamada «coxa genuina,» pero el Meron nunca incluye la región de la articulación trocantéro posterior, y que delimita la ranura es siempre una parte de la sutura basicostal. La coxa con un Meron ampliado tiene una apariencia similar a uno dividido por una sutura coxal caer en línea con la sutura pleural, pero las dos condiciones son fundamentalmente muy diferentes y no se debe confundir. El Meron alcanza el extremo de su salida de la condición habitual en los dípteros. En algunas de las moscas más generalizadas, como en el Tipulidae, el Meron de la pierna del medio aparece como un gran lóbulo de las coxas que sobresale hacia arriba y posteriormente desde la base coxal; en mayores miembros de la orden de que esté totalmente separada de la coxa y forma una placa de la pared lateral del mesotórax.
El trocánter es el segmento basal del telopodite; es siempre un pequeño segmento en la pata de insectos, se puede mover libremente por una bisagra horizontal en la coxa, pero más o menos fija a la base del fémur. Cuando móvil en el fémur de la articulación femoral trochantero es generalmente vertical u oblicua en un plano vertical, dando un ligero movimiento de la producción y la reducción en la unión, aunque solo un músculo reductor está presente. En la Odonata, ambos ninfas y adultos, hay dos segmentos trocantéreo, pero no son móviles el uno del otro; la segunda contiene el músculo reductor del fémur. El segmento trocantéreo solo habitual de insectos, por lo tanto, probablemente representa los dos trocánteres de otros artrópodos fusionados en un segmento aparente, ya que no es probable que la bisagra coxotrochanteral primaria se ha perdido de la pierna. En algunos de los himenópteros una subdivisión basal del fémur simula un segundo trocánter, pero la inserción del músculo reductor en su base atestigua que pertenece al segmento femoral, ya que como se muestra en la pierna del odonato, el reductor tiene su origen en el verdadero segundo trocánter.
El fémur es el tercer segmento de la pata de insectos, es por lo general la parte más larga y más fuerte de la extremidad, pero varía en tamaño desde la enorme fémur posterior saltatorio de Orthoptera a un segmento muy pequeño tal como está presente en muchas formas larvarias. El volumen del fémur está generalmente correlacionado con el tamaño de los músculos tibiales contenidos dentro de él, pero a veces es ampliado y modificado en forma para otros fines que el de acomodar los músculos tibiales. La tibia es característicamente un segmento delgado en insectos adultos, solo un poco más corto que el fémur o el fémur y trocánter combinado. Su extremo proximal forma una cabeza inclinada más o menos clara hacia el fémur, un dispositivo que permite la tibia para ser flexionado cerca contra la superficie inferior del fémur.
El tarso de los insectos corresponde al penúltimo segmento de una extremidad de artrópodos generalizado, que es el segmento de la llama propodito en crustáceos. En insectos adultos se subdivide habitualmente de dos a cinco subsegmentos, o tarsómeros, pero en el Protura, algunos colémbolos, y larvas de insectos holometábolos conserva la forma primitiva de un segmento sencillo. Los subsegmentos del tarso de insectos adultos son por lo general se puede mover libremente sobre sí por membranas que conectan declinadas, pero el tarso no tiene músculos intrínsecos. El tarso de los insectos adultos alados tiene menos de cinco subsegmentos por lo que probablemente se especializó por la pérdida de uno o más subsegmentos o por una fusión de subsegmentos adyacentes. En los tarsos de Acrididae la pieza basal larga evidentemente está compuesta de tres tarsómero unidos, faltando el cuarto y el quinto. El tarsómero basal está a veces notablemente ampliado y se distingue como la tarsómero. En las superficies inferiores de los subsegmentos del tarso en ciertos Orthoptera existen pequeñas almohadillas, el pulvilli tarsal o euplantulae. El tarso es ocasionalmente fusionado con la tibia en larvas de insectos, formando un segmento tibiotarsal; en algunos casos, parece estar eliminado o reducido a un rudimento entre la tibia y el pretarso.
En la mayoría de las especies el fémur y la tibia son los segmentos más largos de la pierna pero las variaciones en las longitudes y robustez de cada segmento se relacionan con sus funciones. Por ejemplo, gressorial y cursorial o caminar y los insectos de tipo de funcionamiento, respectivamente, por lo general tienen fémures y tibias bien desarrollados en todas las patas, mientras que los insectos saltadores como saltamontes han desarrollado de manera desproporcionada fémures y tibias en las patas posteriores. En los escarabajos acuáticos (Coleóptera) y los insectos (Hemíptera), el tibia e y / o tarsos de uno o más pares de piernas por lo general se modifican para la natación (natatorial) con flecos de pelos largos y delgados. Muchos insectos que viven en el suelo, como los grillos topo (Gryllotalpidae), cigarras ninfales (Hemíptera: Cicadidae) y escarabajos (Scarabaeidae), tienen las tibias de las patas delanteras ampliada y modificada para la excavación (fossorial), mientras que las patas delanteras de algunos insectos depredadores, como las crisopas mantíspidas (Neuróptera) y las mantis (Mantodea), están especializadas para atrapar a su presa, o raptorial. La tibia y tarsómero basal de cada pata trasera de las abejas melíferas están modificadas para recolectar y transportar polen.
Abdomen

El abdomen de un insecto adulto consiste típicamente de 11-12 segmentos y está menos esclerotizado que la cabeza o el tórax. Cada segmento del abdomen está representado por un tergo esclerotizado, el esternón, y tal vez una pleurita. El tergo está separado de los esternones adyacentes así como de la pleura por una membrana. Los espiráculos se encuentran en la zona pleural. La variación de esta especie incluye la fusión del tergo y el esternón para formar la dorsal continua o escudos ventrales o un tubo cónico. Algunos insectos tienen un esclerito en la zona pleural llamado laterotergito. Los escleritos ventrales son llamados a veces laterosternitos. Durante la etapa embrionaria de muchos insectos y la etapa postembrionario de insectos primitivos, 11 segmentos abdominales están presentes. En los insectos modernos hay una tendencia hacia la reducción en el número de los segmentos abdominales, pero el número primitivo durante embriogénesis es de 11. La variación en el número de segmentos abdominales es considerable. Si el Apterygota se considerara como indicativos dentro de la especie de los pterygotes, habría una confusión puesto que el adulto Protura tiene 12 segmentos, y los colémbolos tienen 6, por otra parte, la familia de orthopteran acrididae tiene 11 segmentos, y una muestra de fósiles de Zoraptera tiene el abdomen segmentado.
Generalmente, los primeros siete segmentos abdominales de los adultos (los segmentos pregenitales) son similares en estructura y apéndices falta. Sin embargo, los apterygotes (Tisanuros y plateados) y muchos insectos acuáticos inmaduros tienen apéndices abdominales. Los apterygotes poseen un par de estilos; apéndices rudimentarios que son una serie homóloga con la parte distal de las patas torácicas. Y el mesally tiene uno o dos pares conocidos como protrusible (o exsertile), que son vesículas en al menos algunos segmentos abdominales. Estas vesículas se derivan de los inditos coxales y trocánteres (lóbulos interiores anulados) de los apéndices abdominales ancestrales. Las larvas acuáticas y ninfas pueden tener agallas lateralmente en algunos a la mayoría de los segmentos abdominales, pero el resto de los segmentos abdominales consistirá en las partes reproductivas y anales.
La parte anal-genital del abdomen, conocida como la terminalia, consiste generalmente en segmentos de 8 o 9 hasta el ápice abdominal. Los segmentos 8 y 9 tienen los genitales; segmento 10 es visible como un segmento completo en muchos insectos «inferiores» pero siempre carece de apéndices; y el segmento pequeño 11 está representado por un epiprocto dorsal y ventral con un par de paraproctos derivados del esternón. Además poseen un par de apéndices que articulan lateralmente en el segmento de 11 que normalmente se está anillado y con filamentos que se han modificado (por ejemplo, las pinzas de las tijerillas) o reducido en diferentes órdenes de insectos. El filamento caudal anillado y la mediana dorsal del apéndice surgen de la punta del epiprocto en los apterygotes que son la mayoría de las efímeras (Ephemeroptera), y algunos insectos fósiles. Una estructura similar en moscas de piedra ninfales (Plecóptero) es de homología incierta. Estos segmentos abdominales terminales tienen funciones de excreción y sensoriales en todos los insectos, pero en los adultos hay una función reproductiva adicional.
Genitales externos

La sección terminal del abdomen de una mosca escorpión macho se amplía en una «bombilla genital» Panorpa communis
Los órganos que tratan específicamente con el apareamiento y la deposición de huevos se conocen colectivamente como los genitales externos, aunque pueden ser en gran parte interna. Los componentes de los órganos genitales externos de los insectos son muy diversos en forma y con frecuencia tienen un gran valor taxonómico, particularmente entre las especies que aparecen estructuralmente similares en otros aspectos. Los genitales externos masculinos se han utilizado ampliamente para ayudar a distinguir las especies, mientras que los genitales externos femeninos pueden ser más simple y menos variados.
Las terminales de los insectos hembras adultas incluyen estructuras internas para recibir el órgano masculino copulador y sus espermatozoides así como las estructuras externas utilizadas para la ovoposición (puesta de huevos; sección 5.8). La mayoría de los insectos hembra tienen un tubo de puesta de huevos, o ovopositor; sin embargo, está ausente en las termitas, piojos parásitos. Muchos Plecóptero, Ephemeroptera y otros.
Los ovipositores toman dos formas:
- Cierto, o apendicular, formado a partir de apéndices de segmentos abdominales 8 y 9;
- Sustitucional, compuesto por segmentos abdominales posteriores extensibles.
Anatomía interna
Sistema nervioso
El sistema nervioso de un insecto puede ser dividido en un cerebro y un espinal nervioso ventral. La cápsula de la cabeza se compone de seis segmentos fusionados, cada uno con un par de ganglios, o un grupo de células nerviosas fuera del cerebro. Los tres primeros pares de ganglios están fundidos en el cerebro, mientras que los tres siguientes pares se funden en una estructura de tres pares de ganglios bajo del insecto esófago, llamado el ganglio subesophageal.

Los segmentos torácicos tienen un ganglio en cada lado, que están conectados en un par, un par por segmento. Esta disposición también se ve en el abdomen, pero sólo en los primeros ocho segmentos. Muchas especies de insectos han reducido el número de ganglios debido a la fusión o reducción Algunas cucarachas tienen solo seis ganglios en el abdomen, mientras que la avispa Vespa crabro sólo tiene dos en el tórax y tres en el abdomen. Algunos insectos, como la mosca doméstica Musca doméstica, disponen de todos los ganglios del cuerpo fundido en un solo gran ganglio torácico.
Al menos algunos insectos tienen nociceptores, las células que detectan y transmiten sensaciones de dolor. Esto fue descubierto en 2003 por el estudio de la variación en las reacciones de larva de la mosca de la fruta común Drosophila para el toque de una sonda climatizada y otra climatizada. Las larvas reaccionó al contacto de la sonda con calefacción y un comportamiento de giro estereotipada de que no se exhibió cuando las larvas fueron tocados por la sonda sin calefacción A pesar de la nocicepción se ha demostrado en los insectos, no hay un consenso de que los insectos sienten dolor conscientemente.
Aparato digestivo
Un insecto utiliza su sistema digestivo para extraer los nutrientes y otras sustancias de los alimentos que consume La mayor parte de esta comida se ingiere en forma de macromolécula y otras sustancias complejas como proteínas, polisacáridos, grasas, y ácido nucleicos. Estas macromoléculas deben desglosarse por reacción catabólica en moléculas más pequeñas como aminoácidos y azúcares simples, antes de ser utilizada por las células del cuerpo para la energía, el crecimiento o la reproducción. Este proceso de desmontaje se conoce como la digestión. La estructura principal del sistema digestivo de un insecto es un tubo largo cerrado llamado el canal alimentario, que se extiende longitudinalmente a través del cuerpo. El tubo digestivo dirige los alimentos en una dirección: desde la boca hasta el ano. Cuenta con tres secciones, cada una de las cuales realiza un proceso diferente de la digestión. Además del canal alimentario, insectos también tienen un par de glándulas salivales y embalses salivales. Estas estructuras normalmente residen en el tórax, adyacente al intestino anterior. El intestino es donde la casi totalidad de la digestión de los insectos tiene lugar. Se puede dividir en el intestino anterior, intestino medio y intestino posterior.

Intestino anterior
Diagrama estilizado de tubo digestivo del insecto que muestra túbulos de Malpighi, de un insecto del orden Orthoptera.
La primera sección del tubo digestivo es el intestino anterior o estomodeo (elemento 27 en el diagrama numerada). El intestino anterior es la línea con un revestimiento cuticular hecha de quitina y proteínas para la protección de los alimentos duros. El intestino anterior incluye la cavidad bucal (boca), faringe, esófago, y buche y proventrículo(o una parte puede ser altamente modificado), que tanto la comida tienda y significar cuando continúe pasando hacia adelante al intestino medio. Aquí, la digestión comienza como alimento parcialmente masticado es degradado por la saliva de la Glándulas salivares. Como las glándulas salivales producen líquidos y enzima que digiere carbohidratos (en su mayoría amilasas), fuertes músculos en el líquido de la bomba faringe en la cavidad bucal, la lubricación de la comida como el salivarium hace, y ayudando alimentadores de sangre, y xilema y floema alimentadores.

A partir de ahí, la faringe pasa alimentos al esófago, lo que podría ser solo un simple tubo de pasarlo al buche y proventrículo, y luego al intestino medio, como en la mayoría de los insectos. Alternativamente, el intestino anterior podrá ampliarse en un buche muy ampliada y proventrículo, o la cosecha podría ser solo un divertículo, o una estructura llena de líquido, como en algunas especies de Díptera.
Abeja defecando, es importante tener en cuenta la contracción del ano que es lo que proporciona la presión interna.
Intestino Medio
Una vez que los alimentos salen, se pasa al intestino medio (elemento 13 en el diagrama numerado), también conocido como el mesenterón, donde la mayoría de la digestión se lleva a cabo. Proyecciones microscópicas de la pared del intestino medio, llamados microvellosidades, que aumentan el área de superficie de la pared y permiten más nutrientes para ser absorbidos; que tienden a ser cerca del origen del intestino medio. En algunos insectos, el papel de las microvellosidades y dónde están ubicados puede variar. Por ejemplo, microvellosidades especializadas que producen enzimas digestivas pueden más probable es que sea cerca del final del intestino medio, y la absorción de cerca del origen o principio del intestino medio.
Intestino posterior
En el intestino posterior o proctodeo (elemento 16 en el diagrama numerado), las partículas de alimentos no digeridos se unen por ácido úrico para formar bolitas fecales. El recto absorbe el 90% del agua en estos pellets fecales, y el sedimento seco se elimina a continuación, a través del ano (elemento 17), completando el proceso de la digestión. El ácido úrico se forma utilizando los productos de desecho de hemolinfa difundidos desde los tubos de Malpighi (elemento 20). A continuación, se vacía directamente en el canal alimentario, en la unión entre el intestino medio y el intestino posterior. El número de tubos de Malpighi poseído por un insecto dado varía entre las especies, que van desde solo dos túbulos en algunos insectos a más de 100 túbulos en otros.
Sistema respiratorio
El aparato respiratorio de los insectos consiste de un sistema de tubos internos, (tráqueas, y de sacos a través de los cuales circulan los gases ya sea por difusión o por bombeado activo. Los tejidos están expuestos al oxígeno de esta manera (elemento 8 en el diagrama). Dado que el oxígeno es entregado directamente, el sistema circulatorio no se utiliza para transportar el oxígeno, y por lo tanto se reduce enormemente. El sistema circulatorio de insectos no tiene venas o arterias, y en su lugar consiste en poco más que un tubo dorsal único perforada que pulsa peristálticamente. Hacia el tórax, el tubo dorsal (elemento 14) se divide en cámaras y actúa como corazón del insecto. El extremo opuesto del tubo dorsal es como la aorta del insecto circular el hemolinfa, análogo de artrópodos ‘fluido de sangre, dentro de la cavidad del cuerpo. Se lleva el aire a través de aberturas en los lados del abdomen llamado espiráculos.

Hay muchos patrones diferentes de intercambio gaseoso demostrado por diferentes grupos de insectos. Patrones de intercambio de gases en los insectos pueden variar de continuo y difusivo ventilación, a el intercambio de gases discontinua. Durante continua el intercambio de gases, oxígeno se toma en y dióxido de carbono se libera en un ciclo continuo. En el intercambio de gases discontinua, sin embargo, el insecto toma oxígeno mientras está activo y pequeñas cantidades de dióxido de carbono se liberan cuando el insecto está en reposo ventilación difusivo es simplemente una forma de intercambio continuo de gas que se produce por difusión en lugar de tomar físicamente en el oxígeno. Algunas especies de insectos que se sumerge también tienen adaptaciones para ayudar en la respiración. Como larvas, muchos insectos tienen branquias que pueden extraer el oxígeno disuelto en el agua, mientras que otros tienen que subir a la superficie del agua para reponer los suministros de aire, que se celebren o atrapadas en estructuras especiales.
Sistema circulatorio
Sangre de insectos o función principal de la hemolinfa es el de transporte y baña los órganos del cuerpo del insecto. Preparación por lo general menos de 25% del peso corporal de un insecto, que transporta hormonas, nutrientes y desechos y tiene un papel en, la osmorregulación, control de temperatura, inmunidad, de almacenamiento (agua, carbohidratos y grasas) y la función del esqueleto. También juega un papel esencial en el proceso de muda Una función adicional de la hemolinfa en algunos órdenes, puede ser el de la defensa depredador. Puede contener sustancias químicas desagradables y malolientes que actuarán como impedimento a los depredadores hemolinfa contiene moléculas, iones y células que regulan los intercambios químicos entre tejidos, hemolinfa está encerrado en la cavidad del cuerpo del insecto o haemocoel Es. transportados alrededor del cuerpo por combinado de corazón (posterior) y aorta (anterior) pulsaciones, que se encuentran dorsalmente justo debajo de la superficie del cuerpo. Se diferencia de los vertebrados de sangre, ya que no contiene células rojas de la sangre y por lo tanto carece de oxígeno de alta capacidad de carga, y es más similar a la linfa que se encuentra en los vertebrados
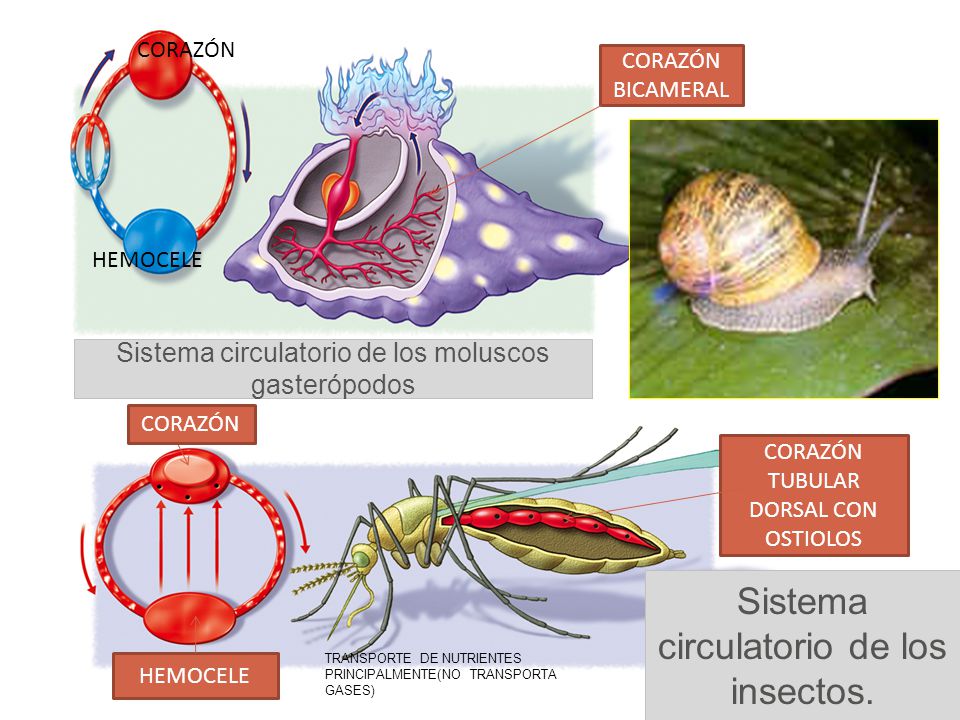
Los fluidos corporales entran a través de una vía Ostia con válvula, que son aberturas situadas a lo largo de la longitud de la aorta combinado y órgano del corazón. Bombeo de la hemolinfa se produce por ondas de contracción peristáltica, originario en el extremo posterior del cuerpo, el bombeo hacia delante en el vaso dorsal, hacia fuera a través de la aorta y luego en la cabeza donde fluye hacia fuera en el hemocele. La hemolinfa se distribuyó a los apéndices de forma unidireccional con la ayuda de las bombas musculares u órganos pulsátiles accesorios que normalmente se encuentran en la base de las antenas o alas ya veces en las piernas, con tasas de bombeo de aceleración con los períodos de mayor actividad. La hemolinfa es particularmente importante para la termorregulación en órdenes tales como Odonata, Lepidoptera, Hymenoptera y Díptera.
Sistema endocrino
Las siguientes glándulas son parte del sistema endocrino
- 1. Células neurosecretoras, en el cerebro.
- 2. Cuerpos cardíacos, detrás del cerebro y a cada lado de la aorta.
- 3. Glándulas protorácicas, un par de glándulas difusas detrás de la cabeza o en el tórax.
- 4. Cuerpo alado (Corpus allatum), par de cuerpos glandulares de origen epitelial, a cada lado del estomodeo.
- Ecdisona
- Hormona juvenil
Aparato reproductor
Hembras
Los insectos hembra son capaces de hacer los huevos, recibir y almacenar el esperma, manipular espermatozoides de diferentes machos, y ponen sus huevos. Sus sistemas reproductivos están formados por un par de ovarios, glándulas accesorias, uno o más espermas, y los conductos que conectan estas partes. Los ovarios producen óvulos y las glándulas accesorias producen las sustancias para ayudar paquete y poner los huevos. Los espermatozoides tienen un recinto donde se producen en diferentes períodos de tiempo y, junto con partes de los oviductos, se puede controlar el uso de esperma. Los conductos y Espermatecas están revestidos con una cutícula.
Los ovarios se componen de un número de tubos, llamados ovariolas, que varían en tamaño y número de cada especie. El número de huevos que el insecto es capaz de hacer varía por el número de ovariolas asimismo la tasa de que los huevos se pueden desarrollar también está influenciado por el diseño ovariole. En los ovarios meroistico, la mayoría de las células hijas se convierten en células de ayuda para un solo ovocito en el clúster. En los ovarios panoistico, cada produce células germinales y del tallo se desarrolla en un ovocito; no hay células auxiliares de la línea germinal. Producción de huevos de los ovarios panoistico tiende a ser más lento que el de ovarios meroistico.
Las glándulas de los oviductos producen una variedad de sustancias para el mantenimiento de esperma, el transporte, y la fertilización, así como para la protección de los huevos. Pueden producir pegamento y sustancias protectoras para el recubrimiento de los huevos o las cubiertas duras para un lote de huevos llamada oteca. Las Espermatecas son tubos o sacos en el que el esperma se puede almacenar entre el momento del apareamiento y el tiempo que un óvulo es fertilizado. La prueba de paternidad de los insectos ha revelado que algunos, y probablemente muchos, insectos hembra utilizan la espermateca y varios conductos para controlar o espermatozoides sesgo utilizado en favor de algunos hombres sobre otros.
Machos
El componente principal del sistema reproductivo masculino son los testículos, suspendido en la cavidad del cuerpo por tráqueas y la grasa corporal. Los insectos apterigotos más primitivos tienen un solo testículo, y en algunos lepidópteros los dos testículos de maduración son secundariamente fusionados en una sola estructura durante las últimas etapas de desarrollo de las larvas, aunque los conductos principales de ellos permanecen separados. Sin embargo, la mayoría de los insectos machos tienen un par de testículos, dentro de los cuales hay tubos de esperma o los folículos que están encerrados dentro de un saco membranoso. Los folículos se conectan a los conductos deferentes por los dos vasos deferentes tubulares conectándose a un conducto eyaculador medial que conduce al exterior. Una porción de los conductos deferentes a menudo se ensancha para formar la vesícula seminal que almacena el esperma antes de que se descargue en la hembra. Las vesículas seminales tienen forros glandulares que secretan nutrientes para la alimentación y el mantenimiento de los espermatozoides. El conducto eyaculador se deriva de una invaginación de las células epidérmicas durante el desarrollo y, como resultado, tiene un revestimiento cuticular. La porción terminal del conducto eyaculador puede esclerotizado para formar el órgano copulador, el edeago. El resto del sistema reproductor masculino se deriva de mesodermo embrionario, a excepción de las células germinales, o espermatogonias, que descienden de las células primordiales de polo muy temprano durante la embriogénesis. El edeago puede ser muy pronunciado o mínimo. La base del edeago puede ser la faloteca parcialmente esclerotizada, también llamada falosoma o teca. En algunas especies la faloteca contiene un espacio, llamado el endosoma (platina de sujeción interno), en el que el extremo de la punta de la edeago puede ser retraída. El conducto deferente es a veces arrastrados a (doblado en) la faloteca junto con una vesícula seminal

Anatomía interna de diferentes taxones
Blattodea
Las cucarachas son más comunes en los climas subtropicales y tropicales. Algunas especies están en estrecha relación con las viviendas humanas y se encuentran frecuentemente alrededor de la basura o en la cocina. Las cucarachas son generalmente omnívoras, con excepción de las cucarachas de la madera, como la Cryptocercus; estas cucarachas son incapaces de digerir celulosa por sí mismas, pero tienen relaciones simbióticas con varios protozoos y bacterias que digieren la celulosa, lo que les permite extraer los nutrientes. La similitud de estos simbiontes en el género Cryptocercus a los de termitas es tal que se ha sugerido que están más estrechamente relacionados con termitass que con otras cucarachas, y la investigación actual apoya firmemente esta hipótesis de las relaciones Todas las especies estudiadas hasta ahora llevan el mutualista obligado endosimbionte bacteriano Blattabacterium, con la excepción de Nocticola australiensis, una especie sin ojos, pigmento o alas que vive en cuevas australianas. Los estudios genéticos recientes indican que son cucarachas muy primitivas
Las cucarachas, como todos los insectos, respiran a través de un sistema de tubos llamados tráqueas Las tráqueas de los insectos se unen a los espiráculos, excepto en la cabeza. Por lo tanto las cucarachas, como todos los insectos, no dependen de la boca y la tráquea para respirar. Los tubos traqueales se ramifican continuamente como un árbol hasta que sus divisiones menores, traqueolas, están asociados con cada célula, permitiendo que el oxígeno gaseoso se disuelva en el citoplasma se extiende a través del revestimiento de la cutícula fina.
Coleóptera
El sistema digestivo de muchos escarabajos está adaptado a la alimentación vegetal; sobre todo es el intestino medio el que realiza la digestión. Sin embargo, en las especies depredadoras (por ejemplo, Carabidae) la mayor parte de la digestión se produce en el buche por medio de las enzimas del intestino medio. En especies de Elateridae, las larvas depredadoras defecan enzimas digestivas sobre su presa, y realizan digestión extraoral. El tubo digestivo comprende básicamente una faringe corta y estrecha, una expansión ampliada, el buche y una molleja poco desarrollada. Después hay un intestino medio, que varía en dimensiones entre las especies, con ciegos o cecum, con longitudes variables. En general, existen de cuatro a seis túbulos de Malpighi.
El sistema nervioso en escarabajos contiene todas las estructuras que se encuentran en los insectos, con variaciones entre las diferentes especies. Con tres ganglios torácicos y siete u ocho ganglios abdominales. Puede ser distinguido a aquel en el que todos ganglios torácicos y abdominales se fusionan para formar una estructura compuesta.
El aparato respiratorio está compuesto de tráqueas que transportan el oxígeno. El aire entra en una serie de tubos a lo largo del cuerpo a través de las aberturas llamadas espiráculos, y es transportado por túbulos cada vez más finos. Los movimientos de bombeo de la fuerza del cuerpo el aire a través del sistema. Algunas especies de escarabajos buceadores llevan una burbuja de aire con ellos cada vez que se sumergen bajo la superficie del agua. Esta burbuja se puede mantener bajo el élitro o puede estar atrapado contra el cuerpo usando pelos especializados. La burbuja por lo general cubre una o más espiráculos por lo que el insecto puede respirar el aire de la burbuja mientras está sumergido. Una burbuja de aire proporciona un insecto con solo un suministro a corto plazo de oxígeno, pero gracias a sus propiedades físicas únicas, el oxígeno se difundirá en la burbuja y desplazando el nitrógeno, llama difusión pasiva, sin embargo, el volumen de la burbuja finalmente disminuye y el escarabajo tendrá que volver a la superficie.
Al igual que otras especies de insectos, los escarabajos tienen hemolinfa en lugar de sangre. El sistema circulatorio abierto del escarabajo es impulsado por un corazón similar a un tubo unido a la parte superior en el interior del tórax.

Diferentes glándulas especializadas para diferentes feromonas producidas para encontrar compañeros. Las feromonas de especies de Rutelinae se producen a partir de células epiteliales que recubre la superficie interna de los segmentos abdominales apicales o feromonas base de aminoácidos de Melolonthinae de las glándulas eversible en el ápice abdominal. Otras especies producen diferentes tipos de feromonas. Dermestidae producir éter, y la especie de Elateridae producen aldehído derivado de ácidos grasos y acetatos. En medio de la búsqueda de un compañero también, luciérnagas (Lampyridae) utilizaron células de grasa corporal modificados woth superficies transparentes respaldadas con cristales de ácido úrico reflectantes para producir biosintéticamente luz, o bioluminiscencia. El producto luz es altamente eficiente, ya que se produce por la oxidación de luciferina por las enzimas luciferasa en la presencia de ATP (trifosfato de adenosina) y oxígeno, produciendo oxiluciferina, el dióxido de carbono, y la luz.
Un notable número de especies han desarrollado glándulas especiales que producen sustancias químicas para disuadir a los depredadores (ver Defensa y la depredación). El Escarabajo de tierra (de Carabidae) tiene glándulas defensivas, situadas en la parte posterior, producen una variedad de hidrocarburos, aldehídos, fenol, quinona, éster, y ácidos liberados de una abertura en el extremo del abdomen. Mientras carábidos africana (por ejemplo, algunas anthia de las cuales utiliza para comprender el género (Thermophilum) emplean las mismas sustancias químicas como las hormigas:. Ácido fórmico
Mientras que el escarabajo bombardero tiene bien desarrollado, al igual que otros escarabajos, glándulas pygidial que vacías de los bordes laterales de las membranas entre segmentos entre el séptimo y el octavo segmento abdominal.
Los órganos auditivos tienen generalmente una membrana (tímpano) exgendida a través de un marco respaldado por un saco de aire y las neuronas sensoriales asociadas. En el orden Coleóptera, órganos timpánicos se han descrito en al menos dos familias. Varias especies del género Cicindela en la familia Cicindelidae tener oídos en la superficie dorsal del primer segmento abdominal debajo del ala; dos tribus de la subfamilia Dynastinae (Scarabaeidae) tienen oídos justo debajo del escudo pronotal o membrana cuello. Las orejas de ambas familias son a frecuencias ultrasónicas, con una fuerte evidencia de que funcionan para detectar la presencia de los murciélagos a través de allí ecolocalización ultrasónica. A pesar de que los escarabajos son un gran pedido y viven en una gran variedad de [nicho ecológico nichos, ejemplos de audiencia es sorprendentemente ausentes en especies, aunque es probable que la mayoría son simplemente sin descubrir.
Dermáptera
El sistema neuroendocrino es típico de los insectos. Tienen un cerebro y un ganglio subesophageal, tres ganglios torácicos, y seis ganglios abdominales. Estos insectos tiene fuertes conexiones neuronales que conectan la corpora neurohemal cardiaca en el cerebro y el ganglio frontal, donde la estrecha relación mediana corpus allatum produce la hormona juvenil en las proximidades de la arota dorsal neurohemal. El sistema digestivo de tijeretas es como todos los otros insectos, que consiste en un primer plano, medio, y el intestino posterior, pero carecen de función gástrica especializada para la digestión en muchas especies de insectos. Los largos y delgados tubos de Malpighi se puede encontrar entre la unión del intestino medio y posterior.

El sistema reproductor de las hembras consiste en un emparejado de ovarios, oviducto lateral, espermateca y una cámara genital. Los conductos laterales están donde los huevos salen del cuerpo, mientras que la espermateca es donde se almacena el esperma. A diferencia de otros insectos, el gonoporo, o la abertura genital están detrás del séptimo segmento abdominal. Los ovarios son órganos primitivos politróficos donde los ovocitos alternan a lo largo de la longitud de la ovariole). En algunas especies de estos largos ovarioles ramifican el conducto lateral, mientras que en otros, ovarioles cortos se encuentran alrededor del conducto.
Díptera
Los genitales de las moscas machos son rotados en un grado variable de la posición que se encuentra en otros insectos. En algunas moscas se trata de una rotación temporal durante el apareamiento, pero en otros es una torsión permanente de los órganos que se produce durante la fase de pupa. Esta torsión puede hacer que el ano esté situado por debajo de los genitales o en el caso de torsión de 360°, que el conducto espermático quede envuelto por el intestino, a pesar de que los órganos externos permanezcan en su posición habitual. Cuando las moscas se aparean, el macho vuela inicialmente sobre la hembra, mirando en la misma dirección, pero luego se da la vuelta para hacer frente en la dirección opuesta. Esto lleva a que las moscas tengan más habilidad de reproducción que la mayoría de los insectos y a un ritmo mucho más rápido. Las moscas vienen en grandes poblaciones habilidad ir debido a aparearse con eficacia y en un corto período de tiempo, especialmente durante la temporada de apareamiento.
La hembra pone sus huevos tan cerca de la fuente de alimento como sea posible, y el desarrollo es muy rápido, lo que permite a la larva consumir comida en un corto período de tiempo antes de transformarse en el adulto. Los huevos eclosionan inmediatamente después de ser despedidos, ciertas moscas son ovovivíparas, la larva eclosiona dentro de la madre. Las larvas de moscas no tienen patas verdaderas, y no poseen demarcación entre el tórax y el abdomen; en las especies más derivadas, la cabeza no se distingue claramente del resto del cuerpo. Los gusanos no tienen extremidades, o bien tienen pequeñas patas. Los ojos y las antenas se reducen o están ausentes, y el abdomen también carece de apéndices tales como cercos. Esta falta de características es una adaptación a un ambiente rico en alimentos, como en materia orgánica en descomposición, o como un endoparásito. La pupa adopta diversas formas, y en algunos casos se desarrollan dentro de un capullo de seda. Después de emerger de la pupa, la mosca adulta rara vez vive más de unos pocos días, y sirve principalmente para reproducir y para dispersarse en busca de nuevas fuentes de alimento.
Lepidóptera
En el sistema reproductivo de las mariposas y las polillas, los órganos sexuales de los machos son complejos y poco claros. En las hembras, hay tres tipos de órganos genitales en función de la taxa relativa: monotrysian, exoporian y dytresian. En el tipo monotrysian hay una abertura en los segmentos fusionados de los esternones 9 y 10, que actúan como la inseminación y la ovoposición. En el tipo exoporian (en los Hepaloidae y Mnesarchaeoidea) hay dos lugares separados para la inseminación y la ovoposición, tanto que se producen en el mismo sterna como el tipo monotrysian, 9/10. En la mayoría de las especies de los genitales están flanqueadas por dos lóbulos suaves, aunque pueden ser especializados y esclerotizados en algunas especies de ovoposición en el área, tales como grietas y tejidos dentro de la planta. Las hormonas y las glándulas que producen ellos producen el desarrollo de las mariposas y polillas por medio del sistema endocrino. La primera hormona de los insectos, es la PTTH (prothoracicotropic hormonal) y opera el ciclo de vida de las especies y la diapausa (ver relaciona la sección) Esta hormona es producida por los corpora allata y los corpora cardiaca, donde también se almacena. Algunas glándulas están especializados para realizar determinada tarea, tales como la producción de la seda o la producción de saliva en los palpos. Mientras los cuerpos cardiaca producir PTTH, el allata corpora también produce hormonas jeuvanile, y las glándulas producen hormonas prothorocic muda.

En el sistema digestivo, la región anterior del intestino anterior se ha modificado para formar una faringe que funciona para succionar ya que de esta manera consumen los alimentos que necesitan y que son en su mayor parte los líquidos. Un esófago conduce posteriormente a la faringe. El intestino medio es corto y recto; el intestino grueso es más largo y enrollado. Los antepasados de las especies de lepidópteros, derivados de los Hymenoptera, tenían el ciego del intestino medio, aunque esto se perdió en las mariposas y las polillas actuales. En lugar de ello, todas las enzimas digestivas distintas de la digestión inicial, se inmovilizan en la superficie de las células del intestino medio. En larvas, de cuello largo y acechado se encuentran en las regiones anteriores y posteriores del intestino medio, respectivamente. En los insectos, las células caliciformes excretan potasio de iones positivos, que son absorbidos a partir de hojas ingeridas por las larvas. La mayoría de las mariposas y polillas muestran el ciclo digestivo normal, sin embargo las especies que tienen una dieta diferente requieren adaptaciones para satisfacer estas nuevas demandas.

En el sistema circulatorio, la hemolinfa o la sangre de insectos, se utiliza para hacer circular el calor en una forma de termorregulación, donde la contracción de los músculos produce calor, que se transfiere al resto del cuerpo cuando las condiciones son desfavorables. En las especies de lepidópteros, la hemolinfa se hace circular a través de las venas de las alas por un órgano pulsante, ya sea por el corazón o por la entrada de aire en la tráquea y se lleva el aire a través de espiráculos a lo largo de los lados del abdomen y el tórax suministro de la tráquea con el oxígeno, ya que pasa a través del sistema respiratorio del lepidóptero. Hay tres diferentes tráqueas que suministran oxígeno difusión de oxígeno a través del cuerpo especies: Las dorsales, ventrales y viscerales. Las tráqueas dorsales suministran de oxígeno a la musculatura dorsal y los vasos, mientras que las tráqueas ventrales suministran a la musculatura ventral y al cordón nervioso mientras que las tráqueas viscerales proporcionan a los intestinos, cuerpos grasos, y gónadas.


