
MORFOLOGÍA VEGETAL
La Morfología vegetal, en sentido amplio, se define como el estudio de la estructura y forma de las plantas, e incluye la Citología y la Histología. La primera se ocupa del estudio fino de la constitución de la célula y la segunda del estudio de los tejidos. Citología e Histología, conjuntamente, son necesarias para comprender la anatomía vegetal, o sea, su constitución interna y, además, son un complemento de la organografía, exomorfología o morfología en sentido estricto, que trata de la forma externa de las plantas.
- Métodos de la morfología vegetal
Las plantas nos ofrecen una infinidad de formas particulares y el objetivo de la morfología es descubrir los patrones o regularidades generales en el fondo de tal diversidad, asimismo comprender y describir tal diversidad desde varios puntos de vista. Para alcanzar este fin se pueden seguir dos caminos:
- La morfología puede estudiarse sobre la base de simples observaciones, sin tener en cuenta criterios de causalidad, mediante el examen comparativo. Mediante suposiciones de muchas formas individuales, en el cual se puede encontrar el plan fundamental o el tipo de un grupo de formas. Este camino es el que sigue la llamada morfología descriptiva y comparativa, que parte de la idea de que la infinidad de formas no son más que variaciones de un mismo tipo de estructura primitiva. Su objeto es reconocer estos tipos y derivarlos de los casos particulares, uno de los principales representantes fue Goethe, quien en su obra Metamorfosis de las plantas (Versuch die Metamorphose der Pflanzen zu erklären) publicada en 1790, trató de encontrar el tipo de la «planta primitiva».
- Es también posible plantearse la cuestión acerca del origen de tales tipos e investigar las causas a las que se deben las concordancias topológicas. Este segundo camino es el de la morfología experimental o analítica, de la que procede la actual fisiología del desarrollo. El objetivo principal del trabajo morfológico en botánica, es la interpretación histórica de las estructuras recientes mediante la consideración de su origen filogenético natural, es decir, comparar la diversidad de formas actuales con las formas comunes iniciales simples, que constituyen, respecto a las actuales, el estado básico o el punto de partida. De este modo el tipo abstracto, de aspecto primitivo, se convierte en la forma ancestral, real, de la cual se ha derivado la actual diversidad de diferenciaciones.
- La citología
La biología celular o bioquímica celular (antiguamente citología, del griego ?????, que significa ‘célula’) es una disciplina académica que se encarga del estudio de las células en cuanto a lo que respecta a las propiedades, estructura, funciones, orgánulos que contienen, su interacción con el ambiente y su ciclo vital. Con la invención del microscopio óptico fue posible observar estructuras nunca antes vistas por el hombre, las células. Esas estructuras se estudiaron más detalladamente con el empleo de técnicas de tinción y de citoquímica y con la ayuda fundamental del microscopio electrónico. La biología celular se centra en la comprensión del funcionamiento de los sistemas celulares, de cómo estas células se regulan y la comprensión del funcionamiento de sus estructuras. Una disciplina afín es la biología molecular.
- Historia
La primera referencia al concepto de célula data del siglo XVII, cuando el inglés Robert Hooke utilizó este término, célula (por su parecido con las habitaciones de los sacerdotes llamadas celdas), para referirse a los pequeños huecos poliédricos que constituían la estructura de ciertos tejidos vegetales como el corcho. No obstante, hasta el siglo XIX no se desarrolla este concepto considerando su estructura interior. Es en este siglo cuando se desarrolla la teoría celular, que reconoce la célula como la unidad básica de estructura y función de todos los seres vivos, idea que constituye desde entonces uno de los pilares de la biología moderna. Fue esta teoría la que desplazó en buena medida las investigaciones biológicas al terreno microscópico, pues las células no son visibles a simple vista. La unidad de medida utilizada es el micrómetro o micra (?m), existiendo células de entre 2 y 20 ?m. La investigación microscópica pronto daría lugar al descubrimiento de la estructura celular interna incluyendo el núcleo, los cromosomas, el aparato de Golgi, las mitocondrias y otros orgánulos celulares, así como la identificación de la relación existente entre la estructura y la función de los orgánulos celulares. Ya en siglo XX, la introducción del microscopio electrónico reveló detalles de la megaestructura celular, y aparecieron la histoquímica y la citoquímica. También se descubrió la base material de la herencia, con los cromosomas y el ADN, y nació la citogenética. Atendiendo a su organización celular, los seres vivos se clasificarían en acelulares (virus, viroides) y celulares, siendo estos últimos a su vez clasificados en eucariotas y procariotas.
- Campos de estudio
Para alcanzar sus objetivos, los biólogos celulares se ven obligados a estudiar los componentes de la célula a nivel molecular (biología molecular).
- Componentes principales del estudio celular:
- membrana plasmática
- La membrana plasmática.
- Membrana celular.
- Membrana citoplasmática o plasmalema, es una bicapa lipídica que delimita todas las células. Es una estructura laminada formada por:
- Fosfolípidos.
- Glicolípidos.
- Proteínas que rodea, limita, da forma y contribuye a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de las células. Regula la entrada y salida de muchas sustancias entre el citoplasma y el medio extracelular. Es similar a las membranas que delimitan los orgánulos de células eucariotas.
- Está compuesta por dos láminas que sirven de «contenedor» para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica.
- Está formada principalmente por fosfolípidos (fosfatidiletanolamina yfosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas).
- La principal característica de esta barrera es su permeabilidad selectiva, lo que le permite seleccionar las moléculas que deben entrar y salir de la célula. De esta forma se mantiene estable el medio intracelular, regulando el paso de agua, ionesy metabolitos, a la vez que mantiene el potencial electroquímico(haciendo que el medio interno esté cargado negativamente). La membrana plasmática es capaz de recibir señales que permiten el ingreso de partículas a su interior.
- Cuando una molécula de gran tamaño atraviesa o es expulsada de la célula y se invagina parte de la membrana plasmática para recubrirlas cuando están en el interior ocurren respectivamente los procesos de endocitosis y exocitosis.
- Tiene un grosor aproximado de 7,5 nm y no es visible al microscopio óptico pero sí al microscopio electrónico, donde se pueden observar dos capas oscuras bilaterales y una central más clara. En las células procariotas y en las eucariotas osmótrofas como plantas y hongos, se sitúa bajo otra capa exterior, denominada pared celular.
- La membrana celular cumple varias funciones: a) delimita y protege las células; b) es una barrera selectivamente permeable, ya que impide el libre intercambio de materiales de un lado a otro, pero al mismo tiempo proporcionan el medio para comunicar un espacio con otro; c) permite el paso o transporte de solutos de un lado a otro de la célula, pues regula el intercambio de sustancias entre el interior y el exterior de la célula siguiendo un gradiente de concentración; d) poseen receptores químicos que se combinan con moléculas específicas que permiten a la membrana recibir señales y responder de manera específica, por ejemplo, inhibiendo o estimulando actividades internas como el inicio de la división celular, la elaboración de más glucógeno, movimiento celular, liberación de calcio de las reservas internas, etc.
- Composición química
Aspectos estructurales de la membrana plasmática y Modelo de mosaico fluido.

- Función. Cada tipo de membrana está especializada en una o varias funciones determinadas dependiendo del compartimento celular donde se encuentre. Entre las múltiples funciones necesarias para la célula que realizan las membranas están la creación de gradientes iónicos, los cuales hacen sensible a la célula frente a estímulos externos, permiten la transmisión de información y la producción de ATP, son necesarios para la realización del transporte selectivo de moléculas, etcétera. Las membranas también hacen posible la creación de compartimentos intracelulares para realizar funciones imprescindibles o la envuelta nuclear que encierra al ADN. En las membranas se disponen múltiples receptores que permiten a la célula «sentir» la información que viaja en forma de moléculas por el medio extracelular, dan a las neuronas sus propiedades y capacidades, también a las musculares. Poseen enzimas asociadas que realizan numerosas actividades metabólicas, como la síntesis de celulosa, fosforilaciones, producción de energía, síntesis de lípidos, etcétera. Como hemos visto en el apartado anterior, la adhesividad celular a la matriz extracelular o a otras células en los tejidos animales se debe a las moléculas presentes en la membrana plasmática.
- Propiedades. Parte de las funciones de las membranas son debidas a sus propiedades físico-químicas: a) Es una estructura fluida que hace que sus moléculas tengan movilidad lateral, como si de una lámina de líquido viscoso se tratase ; b) Es semipermeable, por lo que puede actuar como una barrera selectiva frente a determinadas moléculas; c) Posee la capacidad de ser rota y fusionada de nuevo sin perder su organización, es una estructura flexible y maleable que se adapta a las necesidades de la célula; d) Está en permanente renovación, es decir, eliminación y adición de moléculas que permiten su adaptación a las necesidades fisiológicas de la célula.
- Composición y estructura. Las membranas celulares están formadas por lípidos, proteínas y, en menor medida, por glúcidos. La estructura y la organización de las membranas celulares, así como sus propiedades, están condicionadas fundamentalmente por los lípidos. Éstos son moléculas anfipáticas, con una parte hidrofílica y otra hidrofóbica, que se disponen formando una bicapa lipídica donde las partes hidrofóbicas se encuentran en el centro de la membrana y las hidrofílicas en contacto con el agua. Entre los lípidos se insertan las proteínas denominadas integrales o transmembrana, que poseen secuencias de aminoácidos hidrofóbicos entre las cadenas de los ácidos grasos de los lípidos, y dominios hidrofílicos que están con la solución acuosa intra y extracelula. También hay proteínas asociadas a una u otra superficie de la bicapa lipídica. Los glúcidos no aparecen en todas las membranas, por ejemplo en algunas intracelulares, pero son abundantes en la que delimita la célula con el medio externo, la membrana plasmática, localizados en la superficie extracelular. Los glúcidos se encuentran unidos covalentemente a los lípidos o a las proteínas.
- Membrana celular
Esquema de la organización de una membrana plasmática. Es una bicapa estructurada por los lípidos. Determinados lípidos se asocian entre sí para formar agrupaciones más densas denominadas balsas de lípidos, en la cuales se sitúan ciertas proteínas por afinidad eléctrica. El colesterol se localiza entre las cadenas de ácidos grasos, cerca de la zona hidrofílica («cabezas» de los lípidos). Las proteínas integrales comunican el exterior (arriba) con el interior (abajo) de la célula. Los glúcidos se localizan en la parte extracelular formando el glucocálix. En este esquema no se muestran las interacciones con la matriz extracelular ni con las moléculas del citoesqueleto. (Modificado de Edidin, 2003) Por tanto las membranas son como láminas extensas que cuando se observan en secciones transversales, perpendiculares a sus superficies, con el microscopio electrónico presentan un aspecto trilaminar: dos franjas oscuras que corresponden con las partes hifrofílicas de los lípidos y una franja clara más ancha entre ellas que son sus cadenas de ácidos grasos. A esto se denomina unidad de membrana y es así para todas las membranas celulares. El espesor de las membranas varía entre los 6 y los 10 nm, lo cual indica que no todas las membranas son exactamente iguales.
- Mielina
Vainas de mielina en un nervio periférico. Las propiedades fisiológicas y estructurales de las membranas dependen de la proporción y del tipo de moléculas que las componen: lípidos, proteínas y glúcidos. Así, la membrana de los eritrocitos de rata contiene un 50 % de lípidos, un 40 % de proteínas y un 10 % de glúcidos. Una proporción similar a ésta es la más común entre las membranas plasmáticas de todas las células animales, con algunas excepciones. Por ejemplo, la mielina formada por las membranas plasmáticas de las células de Schwan, que rodean a los axones situados fuera del sistema nervioso central, contienen un 80 % de lípidos y un 20 % de proteínas. Las membranas intracelulares suelen contener una mayor proporción de proteínas que la membrana plasmática. La mayor diferencia la encontramos en las mitocondrias donde el porcentaje de proteínas de su membrana interna llega hasta el 80 %. Por supuesto, lípidos, proteínas y glúcidos son grupos heterogéneos de moléculas y también las membranas celulares se diferencian en la composición y en la proporción de distintos tipos de lípidos, de proteínas y de glúcidos. Además, como dijimos anteriormente, las membranas están en una constante renovación que permite a la célula cambiar su composición. En los siguientes apartados veremos los componentes moleculares, para después tratar las propiedades de las membranas celulares y algunas de sus funciones más importantes. En capítulos posteriores veremos que las membranas celulares de los orgánulos participan de forma determinante en sus funciones, en el trasiego de moléculas en el interior de la célula mediante el denominado tráfico vesicular, así como en la incorporación y liberación de macromoléculas entre el interior y el exterior celular en los procesos de endocitosis y exocitosis, respectivamente.
- Bicapa lipídica
- Componentes lipídicos
El 98 % de los lípidos presentes en las membranas celulares son anfipáticos, es decir que presentan un extremo hidrófilo(que tiene afinidad e interacciona con el agua) y un extremo hidrofóbico (que repele el agua). Los más abundantes son los fosfoglicéridos (fosfolípidos) y los esfingolípidos, que se encuentran en todas las células; le siguen los glucolípidos, así como esteroides (sobre todo colesterol). Estos últimos no existen o son escasos en las membranas plasmáticas de las células procariotas. Existen también grasas neutras, que son lípidos no anfipáticos, pero solo representan un 2 % del total de lípidos de membrana.
- Fosfoglicéridos. Tienen una molécula de glicerol con la que se esterifica un ácido fosfórico y dos ácidos grasos de cadena larga; los principales fosfoglicéridos de membrana son la fosfatidiletanolamina o cefalina, la fosfatidilcolina olecitina, el fosfatidilinositol y la fosfatidilserina.
- Esfingolípidos. Son lípidos de membrana constituidos por ceramida (esfingosina + ácido graso); solo la familia de la esfingomielina posee fósforo; el resto poseen glúcidos y se denominan por ello glucoesfingolípidos o, simplemente glucolípidos. Los cerebrósidos poseen principalmente glucosa, galactosa y sus derivados (como N-acetilglucosamina y N-acetilgalactosamina). Los gangliósidos contienen una o más unidades de ácido N-acetilneuramínico (ácido siálico).
- Colesterol. El colesterol representa un 23 % de los lípidos de membrana. Sus moléculas son pequeñas y más anfipáticas en comparación con otros lípidos. Se dispone con el grupo hidroxilo hacia el exterior de la célula (ya que ese hidroxilo interactúa con el agua). El colesterol es un factor importante en la fluidez y permeabilidad de la membrana ya que ocupa los huecos dejados por otras moléculas. A mayor cantidad de colesterol, menos permeable y más dura es la membrana. Se ha postulado que los lípidos de membrana se podrían encontrar en dos formas: como un líquido bidimensional, y de una forma más estructurada, en particular cuando están unidos a algunas proteínas formando las llamadas balsas lipídicas. Se cree que el colesterol podría tener un papel importante en la organización de estas últimas. Su función en la membrana plasmática es evitar que se adhieran las colas de ácido graso de la bicapa, mejorando la fluidez de la membrana. En las membranas de las células vegetales son más abundantes los fitoesteroles.
- Componentes proteicos
El porcentaje de proteínas oscila entre un 20 % en la mielina de las neuronas y un 70 % en la membrana interna mitocondrial; el 80 % son intrínsecas, mientras que el 20 % restantes son extrínsecas. Las proteínas son responsables de las funciones dinámicas de la membrana, por lo que cada membrana tiene una dotación muy específica de proteínas; las membranas intracelulares tienen una elevada proporción de proteínas debido al elevado número de actividades enzimáticas que albergan. En la membrana las proteínas desempeñan diversas funciones: transportadoras, conectoras (conectan la membrana con la matriz extracelular o con el interior), receptoras (encargadas del reconocimiento celular, adhesión) y enzimas. Las proteínas de la membrana plasmática se pueden clasificar según cómo se dispongan en la bicapa lipídica:
- Proteínas integrales. Embebidas en la bicapa lipídica, atraviesan la membrana una o varias veces, asomando por una o las dos caras (proteínas transmembrana); o bien mediante enlaces covalentes con un lípido o un glúcido de la membrana. Su aislamiento requiere la ruptura de la bicapa.
- Proteínas periféricas. A un lado u otro de la bicapa lipídica, pueden estar unidas débilmente por enlaces no covalentes. Fácilmente separables de la bicapa, sin provocar su ruptura.
- Proteína de membrana fijada a lípidos. Se localiza fuera de la bicapa lipídica, ya sea en la superficie extracelular o intracelular, conectada a los lípidos mediante enlaces covalentes.
En el componente proteico reside la mayor parte de la funcionalidad de la membrana; las diferentes proteínas realizan funciones específicas:
- Proteínas estructurales o de anclaje: estas proteínas hacen de «eslabón clave» uniéndose al citoesqueleto y la matriz extracelular.
- Proteínas receptoras: que se encargan de la recepción y transducción de señales químicas.
- Proteínas de transporte: mantienen un gradiente electroquímico mediante el transporte de membrana de diversos iones.
- Estas a su vez pueden ser:
- Proteínas transportadoras: Son enzimas con centros de reacción que sufren cambios conformacionales.
- Proteínas de canal: Dejan un canal hidrofílico por donde pasan los iones.
- Componentes glucídicos
Están en la membrana unida covalentemente a las proteínas o a los lípidos. Pueden ser polisacáridos u oligosacáridos. Se encuentran en el exterior de la membrana formando el glicocalix. Representan el 8 % del peso seco de la membrana plasmática. Sus principales funciones son dar soporte a la membrana y el reconocimiento celular (colaboran en la identificación de las señales químicas de la célula).
- Funciones
- La función principal de la membrana plasmática es mantener el medio interno separado de la capa fosfolipídica y a las funciones de transporte que desempeñan las proteínas. La combinación de transporte activo y transporte pasivo hacen de la membrana endoplasmático una barrera selectiva que permite a la célula diferenciarse del medio.
- Permite a la célula dividir en secciones los distintos orgánulos y así proteger las reacciones químicas que ocurren en cada uno.
- Crea una barrera selectivamente permeable en donde solo entran o salen las sustancias estrictamente necesarias.
- Transporta sustancias de un lugar de la membrana a otro, ejemplo, acumulando sustancias en lugares específicos de la célula que le puedan servir para su metabolismo.
- Percibe y reacciona ante estímulos provocados por sustancias externas (ligando).
- Mide las interacciones que ocurren entre células internas y externas.
- Poseen receptores químicos que se combinan con moléculas específicas que permiten a la membrana recibir señales y responder de manera específica, por ejemplo, inhibiendo o estimulando actividades internas como el inicio de la división celular, la elaboración de más glucógeno, movimiento celular, liberación de calcio de las reservas internas, etc.
- Diferenciaciones de la membrana
Van dirigidas al desempeño de funciones concretas y consistentes en algún tipo de alteración morfológica del contorno de la célula en cualquiera de sus superficies:
- Superficie apical (que da hacia la luz del conducto): son típicas las microvellosidades de algunas células epiteliales. Se tratan de evaginaciones con forma de dedo de guante que aumentan la superficie de absorción intestinal.
- Superficie basal (lado opuesto a la luz del conducto): también destacan las células epiteliales, concretamente las que en el riñón presentan invaginaciones que aumentan la superficie de reabsorción de agua en el tubo contorneado proximal de las nefronas.
- Superficie lateral: son las denominadas uniones intercelulares que posibilitan las interacciones entre células vecinas. Son de varios tipos: estrechas o impermeables, que no dejan espacio intercelular alguno, comunicantes o en hendidura, que dejan un reducido espacio intercelular, y adherentes o desmosomas, que, aunque con un espacio intercelular mayor, implican una fuerte unión mecánica entre las células.
- Permeabilidad
La permeabilidad de las membranas es la facilidad de las moléculas para atravesarla. Esto depende principalmente de lacarga eléctrica y, en menor medida, de la masa molar de la molécula. Moléculas pequeñas o con carga eléctrica neutra pasan la membrana más fácilmente que elementos cargados eléctricamente y moléculas grandes. Además, la membrana es selectiva, lo que significa que permite la entrada de unas moléculas y restringe la de otras. La bicapa lipídica, debido a su interior hidrofóbico, actúa como una barrera altamente impermeable a la mayoría de moléculas polares, impidiendo que la mayor parte del contenido hidrosoluble de la célula salga de ella. Pero por esta misma razón, las células han tenido que desarrollar sistemas especiales para transporte las moléculas polares a través de sus membranas. Con el tiempo suficiente, esencialmente cualquier molécula difundirá a través de una bicapa lipídica libre de proteínas, a favor de su gradiente de concentración. Sin embargo la velocidad a la que una molécula difunde a través de una bicapa lipídica varía enormemente, dependiendo en gran parte del tamaño de la molécula y de su solubilidad relativa al aceite (es decir, cuanto más hidrofóbica o no polar), tanto más rápidamente difundirá a través de una bicapa. Las moléculas pequeñas no polares se disuelven fácilmente en las bicapas lipídicas y por lo tanto difunden con rapidez a través de ellas. Las moléculas polares sin carga si su tamaño es suficientemente reducido también difunden rápidamente a través de una bicapa. Ejemplos de estas sustancias no polares son los solventes orgánicos, que presentan una polaridad alta o baja. Por ejemplo: el metanol, la acetona, el etanol, la urea, etc. La reacción que provocan en la membrana plasmática, dichos solventes, al no ser capaces de atravesar dicha membrana, es de degradación, al ser moléculas muy polares provocan que la bicapa lipídica se degrade, que sufra un desgaste. Hay que tomar en cuenta que la permeabilidad de cada soluto se expresa como su penetración relativa. Los alcoholes, como ejemplo de ellos el metanol, etanol, butanol, octanol, etc., pueden actuar en las membranas biológicas fundamentalmente de 3 formas:
- alterando la fluidez de las membranas, lo que indirectamente afectaría el funcionamiento de las proteínas como enzimas y canales
- produciendo una deshidratación a nivel de las membranas
- interactuando directamente con las proteínas de la membrana.
La membrana plasmática puede sufrir un proceso llamado lisis, que hace referencia al rompimiento de la membrana, ya sea mecánicamente, químicamente o por alguna combinación de los dos. Para realizar la lisis química, las células se suspenden en una solución que contiene detergentes y otros reactivos que interfieren con los enlaces químicos que sostienen las proteínas de las membranas juntas. Esto resulta en la rotura de la membrana y la liberación de los componentes intracelulares. Existen dos tipos de lisis: la lisis tradicional(mecánica) y la lisis por medio de detergentes(química) haciendo referencia al párrafo anterior: Dentro de la tradicional se encuentran tres ejemplos; Homogenización líquida, donde las células se rompen al ser forzadas a pasar por espacios muy pequeños, Sonificación, aplicada a ondas de alta frecuencia rompen las células y Congelamiento, lo cual son ciclos de congelación continuos rompen la célula induciendo la formación de cristales. De igual manera esta la lisis por medio de detergentes (química), donde los detergentes rompen la barrera lipídica de una manera suave, solubilizando las proteínas e interrumpiendo la interacción lípido-lípido, lípido-proteína y proteína-proteína. Los detergentes, al igual que los lípidos, se asocian entre ellos y se unen a superficies hidrofóbicas. Se componen de una cabeza polar hidrofílica y una cola no polar hidrofóbica. La permeabilidad depende de los siguientes factores:
- Solubilidad en los lípidos: Las sustancias que se disuelven en los lípidos (moléculas hidrófobas, no polares) penetran con facilidad en la membrana dado que está compuesta en su mayor parte por fosfolípidos.
- Tamaño: la más grande parte de las moléculas de gran tamaño no pasan a través de la membrana. Solo un pequeño número de moléculas polares de pequeño tamaño pueden atravesar la capa de fosfolípidos.
- Carga: Las moléculas cargadas y los iones no pueden pasar, en condiciones normales, a través de la membrana. Sin embargo, algunas sustancias cargadas pueden pasar por los canales proteicos o con la ayuda de una proteína transportadora.
También depende de las proteínas de membrana de tipo:
- Canales: algunas proteínas forman canales llenos de agua por donde pueden pasar sustancias polares o cargadas eléctricamente que no atraviesan la capa de fosfolípidos.
- Transportadoras: otras proteínas se unen a la sustancia de un lado de la membrana y la llevan al otro lado donde la liberan.
- Uso del término membrana celular
La expresión membrana celular se usa con dos significados diferentes:
- Membrana plasmática, descrita en el presente artículo, es la membrana que siempre envuelve al citoplasma de las células. Aunque este uso siempre fue ilegítimo, está extraordinariamente extendido, sobre todo en los textos de habla inglesa (cell membrane).
- Pared celular, también llamada membrana de secreción, es una cubierta más o menos resistente que cubre a todas o la mayoría de las células de las plantas, los hongos y los protistas pluricelulares.
- Origen de la ambigüedad
Durante siglo y medio (c.1800-c.1950) la investigación de las células se basó solo en la observación mediante microscopía óptica. Ésta no puede, por razones físicas relacionadas con la longitud de onda de la luz, detectar estructuras de menos de 0,25 µm (micrómetros). Se llamó membrana celular al límite de la célula cuando éste era distinguible, y éste sigue siendo el único uso legítimo de la expresión. En la mayor parte de los casos lo que se observaba era un recubrimiento, más o menos flexible, hecho de polisacáridos, de proteínas o de polímeros mixtos, al que se llama también pared celular. Ésta es precisamente la expresión que debe preferirse para eludir la ambigüedad. A principios del siglo XX, investigaciones experimentales de la fisiología celular condujeron a postular la existencia, en todas las células, de una membrana invisible, a la que se llamó membrana plasmática o citoplasmática, y que debía estar compuesta esencialmente de lípidos. Ésta representaba la envoltura del protoplasma, la parte fisiológicamente activada de la célula. Con el uso del microscopio electrónico, pudo observarse por fin la membrana plasmática, cuyo espesor típico es de solo 0,0075 µm (109 Å).
- citoesqueleto
El citoesqueleto es un orgánulo y también es un entramado tridimensional de proteínas que provee soporte interno en las células, organiza las estructuras internas e interviene en los fenómenos de transporte, tráfico y división celular. En las células eucariotas, consta de filamentos de actina, filamentos intermedios, microtúbulos y septinas, mientras que en las procariotas está constituido principalmente por las proteínas estructurales FtsZ y MreB. Las septinas se consideran el cuarto componente del citoesqueleto. El citoesqueleto es una estructura dinámica que mantiene la forma de la célula, facilita la movilidad celular (usando estructuras como los cilios y los flagelos), y desempeña un importante papel tanto en el tráfico intracelular (por ejemplo, los movimientos de vesículas y orgánulos) y en la división celular. Luego del descubrimiento del citoesqueleto a principios de los años 80 por el biólogo Keith Porter, el Dr. Donald Ingber consideró que, desde un punto de vista mecánico, la célula se comportaba de manera similar a estructuras arquitectónicas denominadas estructuras de tensegridad. La evolución del citoesqueleto ha sido un motivo de estudio actual, a partir de éste enfoque se ha propuesto un modelo de evolución rápida conocido como el modelo de «complejidad temprana». Este modelo propone que a través de procesos de diversificación y especialización de moléculas ancestrales del citoesqueleto (proto-actina y proto-tubulina), se incrementó la complejidad del sistema en el último ancestro común de los eucariontes (LECA, por sus siglas en inglés «last eucaryotic common ancestor»). El incremento de complejidad en el LECA se produjo por un aumento en la cantidad de proteínas que conforman a cada uno de los filamentos, así como por la aparición de un gran número de proteínas motoras y accesorias.
- Mecanismos de movimiento celulares
Todas las células poseen movimientos celulares, como las corrientes citoplasmáticas, los movimientos de los organelos, los cromosomas, y los cambios de morfología celular durante la división. Existen dos mecanismos de movimientos celulares: el montaje de proteínas contráctiles como la actina y la miosina, y las estructuras motoras permanentes formadas por la asociación de microtúbulos (cilios y flagelos). La actina participa en el mantenimiento de la organización citoplasmática, la movilidad celular y el movimiento interno de los contenidos celulares. En algunos casos, el movimiento es producido por la interacción entre actina y miosina, por ejemplo, los movimientos musculares de los vertebrados. Los cilios y flagelos son estructuras largas, delgadas y huecas que se extienden desde la superficie de las células eucariotas. Los cilios son cortos y aparecen en grandes cantidades, los flagelos son largos y escasos. Solo están ausentes en unos pocos grupos de eucariontes (algas rojas, hongos, plantas con flor y gusanos redondos).
- El citoesqueleto eucarióticas
El citoesqueleto es una estructura intracelular compleja importante que determina la forma y el tamaño de las células, así como se le requiere para llevar a cabo los fenómenos de locomoción y división celulares. Además, en el citoesqueleto radica el control del movimiento intracelular de organelos y permite una organización adecuada para que se lleven a cabo los eventos metabólicos requeridos. La estructuración compleja del citoesqueleto está basada en la interacción de un conjunto de proteínas, las cuales se asocian y forman una red intracelular tridimensional. Es la complejidad de la red tridimensional lo que favorece los fenómenos indicados e incluso el que las células tengan la capacidad de adherirse al medio extracelular, así como el que interactúe y se comuniquen entre ellas. Gracias a tales fenómenos, en el citoesqueleto radica la efectividad con la que las células responden al microambiente en el cual se encuentran. El citoesqueleto de las células eucariotas está basado principalmente en tres tipos de filamentos citoesqueléticos: microfilamentos, filamentos intermedios y microtúbulos. Estos filamentos son largos y se polimerizan por cada uno de sus monómeros. Los monómeros de cada filamento están codificados por los genes y las células los sintetizan de forma constitutiva en niveles basales de expresión de proteínas. Sin embargo, la estructura tridimensional que forman los polímeros al constituir el citoesqueleto no está codificada por el material genético y por lo consiguiente, se transmite de una célula a otra por ser parte de su citoplasma. Sin embargo, el citoesqueleto se considera que perdurará siempre por que funciona como una «memoria celular», esto porque es capaz de almacenar e integrar todas las interacciones que se hayan sucedido en él, durante su interacción con el microambiente que se le presente y lo cual influirá en el futuro comportamiento celular como lo indicaron Fletcher y Mullins en el 2010.

- Microfilamentos (actina)
Los microfilamentos tienen un diámetro de unos 5 a 7 nm. Están formadas por una proteína globular llamada actina que puede presentarse de dos formas:
- Actina no polimerizada (G actina): la actina se encuentra asociada a la profilina que evita su polimerización. Representa la mitad de la actina de la célula y es utilizada para polimerizar microfilamentos cuando es necesario.
- Actina polimerizada (F actina): es una doble hélice dextrógira de dos hebras de actina no polimerizada. Esta actina se puede encontrar asociada a otras proteínas:
- Proteínas estructurales: que permiten la unión de los filamentos de actina
- Proteínas reguladoras: la más importante es la miosina que permite la contracción muscular al permitir que la actina se desplace sobre ella.
Las funciones de los microfilamentos de actina es la contracción muscular, la formación de pseudópodos, el mantenimiento de la morfología celular y, en la citocinesis de células animales, forma un anillo contráctil que divide la célula en dos.
- Filamentos intermedios
Son filamentos de proteína fibrosa que van de 8 a 11nm de diámetro, son los componentes del citoesqueleto más estables, dando soporte a los orgánulos (por sus fuertes enlaces), y heterogéneos. Las proteínas que conforman estos filamentos, la citoqueratina, vimentina, neurofilamentos, desmina y la proteína fibrilar acídica de la glía, dependen del tejido en el que se hallen. Su función principal es la organización de la estructura tridimensional interna de la célula (por ejemplo, forman parte de la envuelta nuclear y de los sarcómeros). También participan en algunas uniones intercelulares (desmosomas).
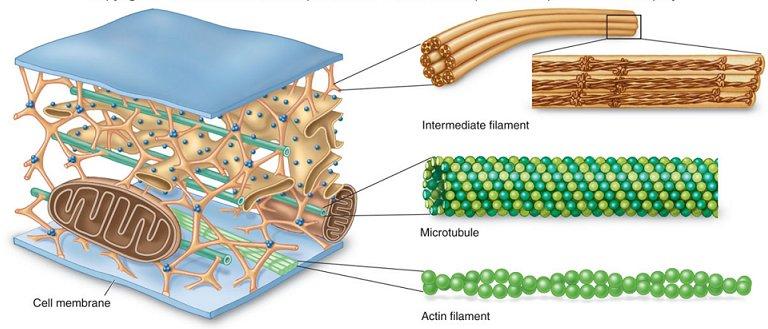
- Microtúbulos
Los microtúbulos son estructuras tubulares de 25 nm de diámetro que se originan en el centro organizador de microtúbulos y que se extienden a lo largo de todo el Citoplasma. Se pueden polimerizar y despolimerizar según las necesidades de la célula. Se hallan en las células eucariotas y están formados por la polimerización de un dímero de dos proteínas globulares, la alfa y la beta tubulina. Cada microtúbulo está compuesto de 13 protofilamentos formados por los dímeros de tubulina. Intervienen en diversos procesos celulares que involucran desplazamiento de vesículas de secreción, movimiento de orgánulos, transporte intracelular de sustancias, así como en la división celular (mitosis y meiosis), ya que forman el huso mitótico. Además, constituyen la estructura interna de los cilios y los flagelos. Los microtúbulos son más flexibles pero más duros que la actina.
- Septinas
Las septinas fueron descritas en levaduras gemantes de Saccharomyces cerevisiae como filamentos en el cuello de la levadura madre, estas observaciones fueron realizadas durante estudios de control genético del ciclo de división celular. Proteínas con secuencias homólogas han sido identificadas en células eucariotas desde levaduras hasta animales incluyendo al hombre, pero no han sido descritas en plantas. En bacterias, las paraseptinas son proteínas relacionadas con las septinas, estas últimas se especula que fueron adquiridas por las células eucariotas por transferencia lateral de bacterias. El número de genes que codifican para septinas varía ampliamente entre los diferentes organismos, por ejemplo para Caenorhabditis elegans se conocen 2, en Encephalitozoon cuniculi 3, en Drosophila melanogaster 5, S. cerevisiae 7,Homo sapiens 14 y en Danio rerio (Pez cebra) 17. Las septinas son un grupo de proteínas que contienen una región conservada de unión a GTP, además todas estas proteínas pueden formar complejos heteroméricos, los cuales se asocian con un alto grado de orden estructural. Por ejemplo, filamentos, anillos y horquillas en forma de reloj de arena los cuales se localizan en el sitio de división de la membrana plasmática, en los anillos de los espermatozoides y en la base de los cilios y dendritas. Las septinas también pueden asociarse con los filamentos de actina, microtúbulos y con fosfolípidos de la membrana. Sin embargo, al igual que los filamentos intermedios, las septinas no presentan polaridad. Aunque se cree que no contribuyen en la generación de fuerza contráctil, se tiene evidencia de su participación en diferentes procesos celulares como la citogénesis, ciliogénesis, neurogénesis, migración celular, mitosis, crecimiento y polarización de las células. Las septinas actúan como andamio y participan en el reclutamiento y organización de proteínas, actuando como barrera de difusión para separar distintos dominios en las membranas, evitando la difusión de factores de la citocinesis, proteínas y RNAs ente la célula madre e hija. El mal funcionamiento de estas proteínas está relacionado con enfermedades neurodegenerativas (Alzheimer y Parkinson),cáncer (colon, oral, leucemias) e infertilidad masculina; por lo cual es importante el estudio de estas proteínas como componentes del citoesqueleto.
- Motores biológicos asociados al citoesqueleto
Existen un grupo de proteínas relacionadas con los distintos elementos del citesqueleto que se especializan en generar movimiento al acoplar sus ciclos de hidrólisis química con ciclos mecánicos de asociación/liberación al filamento. Éstas proteínas motoras (o motores biológicos) amplifican cambios conformacionales provocados por la ruptura del ATP para desplazar orgánelos, vesículas y otras cargas sobre un filamento que les sirve como riel; de éste modo las proteínas motoras participan en una variedad de procesos celulares como la división celular, la migración celular, la endocitosis y la exocitosis por mencionar algunos. Se conocen 3 familias de proteínas motoras: cinecinas, dineínas y miosinas las cuales se desplazan por microtúbulos y micro filamentos respectivamente; hasta el momento no se conocen proteínas motoras que se desplacen por filamentos intermedios y septinas. La interacción entre estos motores biológicos, su desplazamiento a través del citoesqueleto para entregar la carga a su destino final aún no se conoce por completo.
- Cinesina
El supe familia de las cinecinas se constituye por 15 familias las cuales pueden agruparse en 3 grupos dependiendo de la posición del dominio del motor en la secuencia de la molécula. Éstos grupos son las N-cinecinas (dominio de motor en el amino terminal) que se desplazan generalmente hacia el extremo (+) de los microtúbulos, mientras que las C-cinecinas (dominio del motor en el carboxilo terminal) se desplazan hacia el extremo (-), y las M-cinecinas (dominio de motor a la mitad de la molécula) están relacionadas principalmente con la despolimerización de microtúbulos. Algunas cinecinas (de la familia 8 y 14) pueden realizar funciones de desplazamiento y despolimerización.
- Dineínas
Pertenecen a la familia de AAA+ («ATPase associated with diverse activities» por sus siglas en inglés). Las dineínas fueron identificadas por primera vez en 1963 en los cilios de Tetrahymena pyriformis, posteriormente fue aislada en 1987 en tejido cerebral a la que se le denominó dineínas citoplasmática; ésta proteína se desplaza hacia el extremo (-) de los microtúbulos transportando orgánelos como fagosomas, lisosomas, peroxisomas, melanosomas y mitocondrias. Algunos virus como el del VIH, herpesvirus y adenovirus utilizan dineínas citoplasmáticas para transportarse al núcleo. Además las dineínas también participan en funciones relacionadas con la fuerza de tensión de los microtúbulos.
- El citoesqueleto procariota

- FtsZ
FtsZ, una proto-tubulina, fue la primera proteína del citoesqueleto procariota en ser identificada. Al igual que la tubulina, FtsZ forma filamentos en presencia de GTP, pero estos filamentos no se agrupan en microtúbulos. Durante la división celular, FtsZ es la primera proteína que se desplaza al lugar de la división y es esencial para organizar a las proteínas que sintetizan la nueva pared celular en las células que se dividen.
- MreB y ParM
Las proteínas procariotas similares a la actina (también conocidas como proto-actinas), tales como MreB, están involucradas en el mantenimiento de la forma celular. Estas proteínas forman una red helicoidal debajo de la membrana celular que guía a las proteínas que participan en la biosíntesis de la pared celular. Todas las bacterias no esféricas tienen genes que codifican este tipo de proteínas. Algunos plásmidos codifican un sistema de particionado que envuelve una proteína similar a la actina, denominada ParM. Los filamentos de ParM exhiben una inestabilidad dinámica y pueden particionar los plásmidos de ADN durante la división celular en un mecanismo análogo al utilizado por los microtúbulos durante mitosis de los eucariotas.
- Crescentina
La bacteria Caulobacter crescentus contiene una tercera proteína, llamada crescentina, que está relacionada con los filamentos intermedios de las células eucarióticas. La crescentina también participa en el mantenimiento de la forma celular, pero el mecanismo actualmente es poco claro.
- Proteínas WACA
Las proteínas WACA pertenecen a la familia de ATPasas, presentan variación del motivo Walker A (KXXXXGKT), y están ampliamente distribuidas en los procariontes. En la mayoría de las bacterias los genes asociados codifican para uno o más miembros de estas proteínas y las cuales incluyen a las proteínas ParA, MinD, Soj, SopA, Parf, IncC y, probablemente, MipZ. MinD está involucrada en los procesos de división celular, su dinámica varía de acuerdo al organismo, por ejemplo en Escherichia coli se mueve de un extremo de la célula a otro, mientras que en Bacillus subtilis se mantiene en los polos de la célula. Para y Soj, participan en los procesos de segregación de cromosomas, transcripción y organización de plásmidos. En general el papel específico de esta familia de proteínas y su mecanismo molecular son poco entendidos. El citoesqueleto, la tensegridad y la mecanotransducción celular El citoesqueleto es dinámico y no por ello pierde la capacidad del mantenimiento de la forma, la funcionalidad y la estructura de la red tridimensional que lo conforma. Uno de los sitios más recomendables de la WEB para observar mediante visualización científica lo que se ha generado al respecto, y para el cual se aplica el conocimiento generado al momento para el interior de una célula y su relación con la membrana plasmática, es el sitio de XVIVO. En este sitio, en «The inner life of the cell» se puede observar lo que podría suceder al interior de unas células y la relación que con ello tiene el citoesqueleto, el cual está sujeta a propiedades biomecánicas relacionadas con tensión y compresión, las cuales son medibles y explicables mediante las leyes de la física relacionadas con la biomecánica. El balance entre estas propiedades le confieren a la célula una integridad tensional (conocida en el idioma inglés como «Tensegrity») y la cual se basa en lo visualizado en 1993 por el Dr. Donald Ingber, científico que trasladó el concepto arquitectónico (en el cual se le conoce como tensegridad) al ámbito intracelular y que se mantiene vigente en nuestros días. En este sentido, una forma de ampliar visualmente la influencia de los fenómenos de tensión, longitud, rigidez, compresión producidas por las proteínas del citoesqueleto actina y tubulina, así como de la matriz extracelular y las integrinas, es lo presentado en la página WEB del Children’s Hospital Boston denominado «Tensegrity in a Cell»; sitio en el cual las animaciones producidas de manera interactiva por la influencia de las fuerzas indicadas generan cambios en las células y los cuales pueden ser comparados con imágenes obtenidas mediante el microscopio de fluorescencia. La estructuración y la dinámica del citoesqueleto dependen de la forma en que la célula se relaciona con la matriz extracelular y tal relación es lo que determina la biomecánica de las células. Un ejemplo de ello podría ser la dinámica con la que las células ciliadas se presentan ante su entorno como lo propuesto para las células flama de los protonefridios del cestodo Taenia solium. Recientemente, Hersen y Ladoux han hecho referencia a que el mecano biología es un campo emergente que investiga como las células vivas sienten y responden a las fuerzas mecánicas de su entorno. Su comentario hace referencia a que las células están continuamente percatándose de las fuerzas que se suceden a su alrededor aun cuando se encuentran en migración. Tales fuerzas inducen que las células no solo sufran deformaciones sino que también inducen a que se presenten fenómenos como señalización por adhesión y reorganización del citoesqueleto. Estos fenómenos, en referencia a la estrategia experimental que publicaron Delanoë-Ayari y colaboradores, indican que una célula tiene la capacidad de sentir tanto las fuerzas horizontales como las verticales que se presentan durante su desplazamiento y que muestran la importancia que juega la interacción tridimensional entre las células y la matriz extracelular. Las características mecánicas de la matriz extracelular (rigidez y deformabilidad) son factores importantes que influyen en la conducta y la dinámica de las células tales como la diferenciación, la proliferación, la supervivencia, la polaridad y la migración. La mecanotransducción, que se ha establecido como la transformación de fuerzas físicas en señales químicas, es capaz de generar una morfogénesis de un epitelio y ello se puede dar por la generación de modificaciones postransduccionales como la fosforilación de filamentos intermedios como lo demostrado recientemente con el estudio del nematodo Caenorhabditis elegans. Esto resulta un aspecto interesante de la dinámica de la reestructuración del citoesqueleto, ya que se ha encontrado que con los estudios que se efectuaron se muestran que los filamentos intermedios también se mueven y no solo son de soporte y estructura celular. Esto abre un universo importante de como en un ambiente tisular las células contráctiles pueden ejercer influencia en las células de epitelio para que se diferencien y con ello, se favorezcan aspectos de regeneración tisular o diseminación de procesos cancerosos. La tensión con que se presenta el citoesqueleto de una célula, en un momento dado, está influenciado por la dinámica celular y la forma de su núcleo. Cualquier aspecto que induzca cambios en las fuerzas intracelulares que ejercen los componentes del citoesqueleto, derivados de su interacción con el medio extracelular, induce a que también se den cambios en la forma de los núcleos celulares. La constitución del núcleo celular, relacionada con su viscoelasticidad, puede tener un papel determinante en las interacciones biomecánicas que se dan entre el núcleo, el citoesqueleto y la matriz extracelular. Además, sus propiedades visco elásticas podrían tener importantes implicaciones en el estudio de la transducción de señales mecánicas. Se sabe que el núcleo tiene comportamiento como un sólido viscoelástico y por ello presenta propiedades distintas a las del citoplasma. Por lo consiguiente, es de esperarse que cualquier deformación que sufra, así como las propiedades mecánicas que presenta núcleo pudieran estar influenciadas por el estado de tensión-compresión al que esté sometida una célula. Los núcleos celulares también tienen una dinámica propia debida a su composición; cuando una célula va de un lado a otro o bien, pasa a través de un diámetro menor al suyo, la deformación del núcleo también se presenta acorde al que presenta la célula completa. El tamaño y la forma de los núcleos celulares es variable y depende del tipo celular. Su dinámica está asociada a la del citoesqueleto y por lo consiguiente, la composición del núcleo esqueleto está intrínsecamente conectada al citoesqueleto. De hecho se ha indicado que la plasticidad del núcleo celular en las células cancerosas es una determinante para que éstas se diseminen Mecanotransducción es un término que implica que las fuerzas mecánicas aplicadas a las células se transforman en sucesos bioquímicos relevantes y que debido a ellos, se generan diferentes procesos asociados al desarrollo, la fisiología y la patología. Según los autores de la revisión citada, la mecanotransducción celular es un campo de estudio de rápido avance en la investigación científica actual. La relación que el citoesqueleto guarda con la mecanotransducción es estrecha: las células son materiales deformables que basan su forma y tamaño en el citoesqueleto y por lo consiguiente, cualquier fuerza que las afecte genera cambios que se traducen en distintas actividades celulares. Según lo que se ha establecido, son las propiedades de viscoelasticidad del citoesqueleto lo que define sus propiedades mecánicas y que gracias a ellas tenga la plasticidad requerida. Sin embargo, esto es un tema aún controversial. La demostración de cómo se presenta la mecano transducción es un reto tecnológico muy interesante ya que son varias las formas con que este fenómeno puede ser evaluado a nivel de las células y van desde la compresión de membranas, el corte por estrés, el uso de pinzas ópticas, la aplicación de fuerzas magnéticas, el uso del microscopio de fuerza atómica y la aspiración con micro pipetas entre otras
- Papel de la mecano transducción en la invasión por patógenos
Según Hoffman et al el prendido y apagado del mecano transducción (del inglés «switch-like model») está integrado por tres fenómenos: el mecano sensación, el mecano transmisión y el mecano respuesta. Cuando las células responden a estímulos mecánicos tanto externos como internos, un conjunto molecular denominado mecano sensor sufre cambios conformacionales que le permite a las células el detectar tales estímulos. Luego estos estímulos son transmitidos al interior celular a través de los largos filamentos del citoesqueleto y esto se refiere al fenómeno de mecano transmisión. Las señales generadas por estos estímulos se transducen en la activación de señales intracelulares en las que participan segundos mensajeros, con lo que finalmente se genera una mecano respuesta celular. Hay que considerar que estos fenómenos se presentan de forma secuencial en un intervalo de tiempo del orden de cientos de milisegundos y que pueden ser acelerados o retrasados por cambios en la intensidad y la frecuencia de las fuerzas, así como de las condiciones del microambiente que los originaron. Un ejemplo de la importancia de la mecano transducción en la invasión por patógenos es la propuesta para hongos como Cándida albicans y Magnaporthe grisea hecha por Kumamoto en 2008. Un trabajo muy interesante en el que se induce una fuerza magnética a trofozoítos de la ameba Entamoeba histolytica, los cuales previamente se activaron por la fagocitosis de perlitas magnéticas recubiertas con proteínas humanas séricas, muestra que la mecano transducción generada induce a que la célula modifique su migración hacia un solo sitio y que la mecano sensación se da por activación de la cinasa de fosfatidilinositol y la reestructuración de la actina. Según estos autores, la mecano transducción inducida en estos patógenos, que orienta la dirección de migración de ellos, podría estar relacionado con cambios en su virulencia, lo cual podría ser determinante en la invasión de los tejidos humanos infectados. La mecano transducción también podría ser relevante para el comportamiento de los patógenos durante su interacción con las células de sus hospederos; recientemente, mediante la evaluación de la adhesión de protozoarios de Gardia lamblia a vidrio por marcaje fluorescente in vivo del disco suctor y de los flagelos ventrales, se encontró que la fuerza con la que el disco suctor de estos parásitos se adhiere a su sustrato es tan fuerte, lo cual podría ser suficiente para evitar que el parásito se despegue. Esta estructura celular de G. lamblia tiene un diseño tal como una copa de succión que genera un vacío gracias al cual se adhiere firmemente al tejido intestinal. Esto si se tradujese a lo que el parásito hace con el epitelio intestinal, podría dar una explicación de porqué aun cuando el intestino entre en peristalsis, este tipo de protozoarios permanezcan adheridos.
- El citoesqueleto durante la migración celular
La migración celular es un término usado para referirse a fenómenos que implican el desplazamiento de las células, lo cual puede ocurrir en diferentes sustratos; por ejemplo, el suelo en el caso de amibas como Naegleria fowleri, bajo condiciones in vitro o bajo condiciones in vivo (dentro de los organismos). La migración es una respuesta a diferentes estímulos como la necesidad de alimentarse de las células, cambios morfológicos (embriogénesis, organogénesis y regeneración de heridas) o, bien, ante la presencia de factores solubles que estimulan y señalizan a otros eventos tales como la inflamación. Según el contexto en el que las células migran y el tipo de célula involucrada, existen diferentes formas de migración. Una de las más conocidas, dependiente de Quimioatrayentes, es llamada Quimiotaxis. A diferencia de la quimiotaxis, la Quimioquinesis genera un desplazamiento azaroso. La migración celular en respuesta a un ligando que se encuentra unido o inmóvil a una matriz se denomina Haptotaxis. Si la migración depende del sustrato (en el que es importante la topografía de la superficie, su naturaleza química, su rugosidad, etc.) se presenta una adhesión y se activan diversos mecanismos de interacciones moleculares (integrinas, cinasa de adhesión focal) y se genera una reorganización del citoesqueleto y ello ha generado un fenómeno denominado durotaxis. La durotaxis es la tendencia de las células para avanzar hacia sustratos más rígidos (por ejemplo metales como cobre), pero no ha sido completamente descrita. Se sabe que, en caso de células mesenquimales humanas, participan integrinas, cinasa de adhesión focal (FAK) y miosinas no musculares tipo II. Es interesante que durante el fenómeno en que se presenta la duro taxis, hay un cambio de señales mecánicas a bioquímicas (mecano transducción), por lo cual se les debe tomar en cuenta cuando se intente el reproducir in vitro un fenómeno biológico o bien, cuando se busque efectuar una terapia celular. La dinámica del citoesqueleto es crucial para que las células vayan de un lugar a otro como se ilustra con la serie de imágenes y videos obtenidos experimentalmente bajo la excelente composición interactiva concebida por el Dr. Vic Small y que se ha denominado como un viaje visual de la motilidad celular. En este sitio uno puede percatarse de lo interesante que resultan tanto la forma como el tamaño que adoptan las células en un momento determinado durante su migración y que aun así de haber desplegado tal dinámica y reorganización, las células no pierden la capacidad de regresar a su estado original cuando éstas se encuentran en reposo. Aun así, el citoesqueleto en la célula en reposo es dinámico, no se detiene porque son perennes las funciones básicas de tráfico y movimiento intracelulares. Como ya había sido descrito anteriormente, al hacer referencia a la vida interior de las células y lo que de manera animada se presenta en el sitio; una célula se desplaza de un lugar a otro, interacciona con otras células y durante estos fenómenos puede cambiar radicalmente su forma y tamaño pero no deja de tener una dinámica intracelular que le ofrece el citoesqueleto. Un excelente ejemplo de la migración celular inducida por sustancias que atraen células y que provienen de otras dañadas, con fines de reparación de estas últimas, es la migración de neutrófilos luego de su adhesión desde los sinusoides hepáticos hacia los focos de hepatocitos dañados durante el fenómeno de inflamación estéril y del cual se puede observar un interesante video en la sección VideoLab de la revista Science. En el video, los neutrófilos teñidos con fluorescencia en color verde, sufren modificaciones en su forma y tamaño durante su migración hacia el foco de hepatocitos dañados (teñidos fluorescentemente en color rojo) a los cuales intentan restaurar. Previo a su migración, los neutrófilos se encuentran adheridos a las paredes de las sinusoides hepáticas (teñidas fluorescentemente en color azul) y cambian su forma y tamaño al dirigirse hacia el foco mencionado. Con la tecnología microscópica actual es posible observar y videofilmar la manera en que el citoesqueleto se reestructura durante la migración celular. Los recursos tecnológicos son diversos y ellos permiten la visualización desde el nivel micrométrico hasta el nanométrico. Las necesidades de conocer que eventos se suceden en el interior de una célula durante su migración es una preocupación que, por su estudio, se espera que puedan ser mejorados otros aspectos de la biología celular pocos conocidos. Un ejemplo de ello es la suma de esfuerzos de investigadores que estudian la migración celular. Debido a los estudios que se han realizado en células que migran, se ha demostrado que ellas se desplazan mediante la continua interacción con la matriz extracelular que les rodea mediante la interacción continua con focos de adhesión o puntos focales. La forma en que las células interaccionan con dicha matriz, depende de la composición y forma de la misma, por lo consiguiente las células adoptan la forma del medio en el que se encuentran desplazando, como se demostró mediante video microscopía y el uso de marcadores fluorescentes (Doyle et al, 2009). El material suplementario asociado al trabajo de estos autores, es una muestra fantástica de como las células adquieren tal migración e incluso se puede observar al mismo tiempo (ver video 4 en relación a la migración de queratinocitos) como realizan sus movimientos intracelulares el citol esqueleto pertenece a las células vegetales y animales. La migración in vivo de células dendríticas puede ser visualizada por medio de marcadores como 19F/1H usando imagen por resonancia magnética nuclear en 3D en ratones. La técnica puede ser vista con más detalle en la página web de la revista JoVE (por sus siglas en inglés The Journal of Visualized Experiments); la cual da paso a paso la técnica y detalles de ella.
- núcleo celular

- Envoltura nuclear
- Ribosomas
- Poros nucleares
- Nucléolo
- Cromatina
- Núcleo celular
- Retículo endoplasmático
- Nucleoplasma
En biología, el núcleo celular es un orgánulo membranoso que se encuentra en el centro de las células eucariotas. Contiene la mayor parte del material genético celular, organizado en múltiples moléculas lineales de ADN de gran longitud formando complejos con una gran variedad de proteínas como las histonas para formar los cromosomas. El conjunto de genes de esos cromosomas se denomina genoma nuclear. La función del núcleo es mantener la integridad de esos genes y controlar las actividades celulares regulando la expresión génica. Por ello se dice que el núcleo es el centro de control de la célula. La principal estructura que constituye el núcleo es la envoltura nuclear, una doble membrana que rodea completamente al orgánulo y separa ese contenido del citoplasma, además de contar con poros nucleares que permiten el paso a través de la membrana para la expresión genética y el mantenimiento cromosómico. Aunque el interior del núcleo no contiene ningún subcompartimento membranoso, su contenido no es uniforme, existiendo una cierta cantidad de cuerpos subnucleares compuestos por tipos exclusivos de proteínas, moléculas de ARN y segmentos particulares de los cromosomas. El mejor conocido de todos ellos es el nucléolo, que principalmente está implicado en la síntesis de los ribosomas. Tras ser producidos en el nucléolo, éstos se exportan al citoplasma, donde traducen el ARNm.
- Historia
La descripción conocida más antigua de las células y su núcleo por Anton van Leeuwenhoek en 1719. Dibujo de una glándula salival de Chironomus realizado por Walther Flemming en 1882. El núcleo contiene cromosomas politénicos. El núcleo fue el primer orgánulo en ser descubierto. Probablemente, el dibujo más antiguo que se conserva de este orgánulo se remonta a uno de los primeros microscopistas, Anton van Leeuwenhoek(1632-1723). Este investigador observó un hueco o «lumen», el núcleo, en eritrocitos de salmón. Al contrario que los eritrocitos de mamífero, los del resto de vertebrados son nucleados. El núcleo también fue descrito en 1804 por Franz Bauer, y posteriormente con más detalle por el botánico escocés Robert Brown en una charla dictada ante la Sociedad linneana de Londres en 1831. Brown estaba estudiando la estructura microscópica de las orquídeas cuando observó un área opaca, que llamó areola o núcleo, en las células de la capa externa de la flor, si bien no sugirió una función potencial para tal estructura. En 1838 Matthias Schleiden propuso que el núcleo desempeñaba un papel en la generación de células, denominándolo por ello «citoblasto» (constructor de células). Pensaba que había observado células nuevas alrededor de estos «citoblastos». Franz Meyen fue un fuerte opositor de esta opinión habiendo descrito previamente células que se multiplicaban por división y creyendo que muchas células carecerían de núcleo. La idea de que las células se podían generar de novo, bien por el «citoblasto» o bien de otro modo, contradecía los trabajos de Robert Remak (1852) y Rudolf Virchow (1855) quienes propagaron decisivamente el nuevo paradigma de que las células solo eran generadas por otras células («Omnis cellula e cellula»). La función del núcleo permanecía sin aclarar. Entre 1876 y 1878 Oscar Hertwig publicó varios estudios sobre la fecundación de huevos de erizo de mar, mostrando que el núcleo del espermatozoide entraba en el oocito, fusionándose con su núcleo. Esta fue la primera vez que se sugirió que un individuo se desarrollaba a partir de una sola célula nucleada. Esto estaba en contradicción con la teoría de Ernst Haeckel que enunciaba que se repetía la filogenia completa de una especie durante el desarrollo embrionario, incluyendo la generación de la primera célula nucleada a partir de una «monerula», una masa desestructurada de moco primordial («Urschleim», en alemán). Por tanto, la necesidad del núcleo espermático para la fecundación estuvo en discusión por un tiempo. No obstante, Hertwig confirmó su observación en otros grupos animales, como por ejemplo en anfibios y moluscos. Eduard Strasburger obtuvo los mismos resultados en plantas (1884). Esto allanó el camino para la asignación de un papel importante del núcleo en la herencia. En 1873 August Weismann postuló la equivalencia de las células germinales paternas y maternas en la herencia. La función del núcleo como portador de información genética se hizo patente solo después, tras el descubrimiento de la mitosis y el redescubrimiento de la herencia mendeliana a principios del siglo XX. Esto supuso el desarrollo de la teoría cromosómica de la herencia.
- Estructuras
El núcleo es el orgánulo de mayor tamaño en las células animales. En las células de mamífero, el diámetro promedio del núcleo es de aproximadamente 6 micrómetros (?m), lo cual ocupa aproximadamente el 10 % del total del volumen celular. En los vegetales, el núcleo generalmente presenta entre 5 a 25 µm y es visible con microscopio óptico. En los hongos se han observado casos de especies con núcleos muy pequeños, de alrededor de 0,5 µm, los cuales son visibles solamente con microscopio electrónico. En las oósferas de Cycas y de coníferas alcanza un tamaño de 0,6 mm, es decir que resulta visible a simple vista. El líquido viscoso de su interior se denomina nucleoplasma y su composición es similar a la que se encuentra en el citosol del exterior del núcleo. A grandes rasgos tiene el aspecto de un orgánulo denso y esférico.
- Envoltura y poros nucleares
- Lámina nuclear
En las células animales existen dos redes de filamentos intermedios que proporcionan soporte mecánico al núcleo: la lámina nuclear forma una trama organizada en la cara interna de la envoltura, mientras que en la cara externa este soporte es menos organizado. Ambas redes de filamentos intermedios también sirven de lugar de anclaje para los cromosomas y los poros nucleares. La lámina nuclear está compuesta por proteínas que se denominan proteínas laminares. Como todas las proteínas, éstas son sintetizadas en el citoplasma y más tarde se transportan al interior del núcleo, donde se ensamblan antes de incorporarse a la red preexistente. Las láminas también se encuentran en el interior del nucleoplasma donde forman otra estructura regular conocida como velo nucleoplásmico, que es visible usando interface. Las estructuras de las láminas que forman el velo se unen a la cromatina y mediante la disrupción de su estructura inhiben la transcripción de genes que codifican para proteínas. Como los componentes de otros filamentos intermedios, los monómeros de lámina contienen un dominio alfa hélice utilizada por dos monómeros para enroscarse el uno con el otro, formando un dímero con un motivo en hélice arrollada. Dos de esas estructuras dimétricas se unen posteriormente lado con lado dispuestos de modo antiparalelo para formar un tetrámero denominado protofilamento. Ocho de esos protofilamentos se disponen lateralmente para formar un filamento. Esos filamentos se pueden ensamblar o desensamblar de modo dinámico, lo que significa que los cambios en la longitud del filamento dependen de las tasas en competición de adición y desplazamiento. Las mutaciones en los genes de las láminas conducen a defectos en el ensamblaje de los filamentos conocidas como laminopatías. De éstas, la más destacable es la familia de enfermedades conocida como progerias, que dan la apariencia de un envejecimiento prematuro a quienes la sufren. Se desconoce el mecanismo exacto por el que los cambios bioquímicos asociados dan lugar al fenotipo progeroide.
- Cromosomas
Un núcleo celular de fibroblasto de ratón en el que el ADN está teñido de azul. Los diferentes territorios del cromosoma 2 (rojo) y cromosoma 9 (verde) están teñidos mediante hibridación fluorescente in situ. El núcleo celular contiene la mayor parte del material genético celular en forma de múltiples moléculas lineales de ADN conocidas como cromatina, y durante la división celular ésta aparece en la forma bien definida que se conoce como cromosoma. Una pequeña fracción de los genes se sitúa en otros orgánulos, como las mitocondrias o los cloroplastos de las células vegetales. Existen dos tipos de cromatina: la eucromatina es la forma de ADN menos compacta, y contiene genes que son frecuentemente expresados por la célula. El otro tipo, conocido como heterocromatina, es la forma más compacta, y contiene ADN que se transcribe de forma infrecuente. Esta estructura se clasifica a su vez en heterocromatina facultativa, que consiste en genes que están organizados como heterocromatina solo en ciertos tipos celulares o en ciertos estadios del desarrollo, y heterocromatina constitutiva, que consiste en componentes estructurales del cromosoma como los telómeros y los centrómeros. Durante la interface la cromatina se organiza en territorios individuales discretos, los territorios cromosómicos. Los genes activos, que se encuentran generalmente en la región eucromática del cromosoma, tienden a localizarse en las fronteras de los territorios cromosómicos. Se han asociado anticuerpos a ciertos tipos de organización cromatínica, en particular los nucleosomas con varias enfermedades autoinmunes como el lupus eritematoso sistémico. Estos son conocidos como anticuerpos antinucleares (ANA) y también se han observado en concierto con la esclerosis múltiple en el contexto de una disfunción inmune generalizada. Como el caso antes mencionado de la progeria, el papel que desempeñan los anticuerpos en la inducción de los síntomas de la enfermedad autoinmune no está todavía aclarado.
- Nucléolo

- Otros cuerpos subnucleares

- Tamaño de la estructura subnuclear
| Nombre de la estructura | Diámetro de la estructura |
|---|---|
| Cuerpos de Cajal | 0,2-2,0 µm |
| PIKA | 5 µm |
| Cuerpos PML | 0,2-1,0 µm |
| Paraspeckles | 0,2-1,0 µm |
| Speckles | 20-25 nm |
Además del nucléolo, el núcleo contiene una cierta cantidad de cuerpos delimitados no membranosos. Entre éstos se encuentran los cuerpos de Cajal(cuerpos enrollados), los llamados «Géminis de los cuerpos enrollados» (Gemini of coiled bodies, en inglés), la denominada Asociación Cariosómica Polimórfica Interfásica (PIKA, por sus siglas en inglés de Polymorphic Interphase Karyosomal Association), los Cuerpos de la Leucemia Promielocítica (PMLs, por sus siglas en inglés de promyelocytic leukaemia), los «paraspeckles» y los «specles de ayuste» o «motas de empalme» («splicing speckles» en inglés). Aunque se sabe poco sobre el número de estos dominios subnucleares, son significativos en cuanto que muestran que el nucleoplasma no es una mezcla uniforme, sino que más bien contiene subdominios funcionales organizados. Otras estructuras subnucleares aparecen como parte de procesos patológicos. Por ejemplo, se ha visto la presencia de pequeños bastones intranucleares en algunos casos de miopatía nemalínica. Esta enfermedad se produce típicamente por mutaciones en el gen de la actina, y los bastones en sí mismos están constituidos por la actina producida a partir de tales genes mutantes, así como otras proteínas del citoesqueleto.
- Cuerpos de Cajal y GEMs
El núcleo típico posee de 1 a 10 estructuras compactas denominadas Cuerpos de Cajal o cuerpos enrollados (CBs, por sus siglas en inglés de Coiled Bodies), cuyo diámetro mide entre 0,2 µm y 2,0 µm dependiendo del tipo celular y especie. Cuando se observan bajo el microscopio electrónico, se asemejan a ovillos de hilos enmarañados, y son focos densos de distribución de la proteína coilina. Los CBs están implicados en varios tipos distintos de funciones relacionadas con el procesamiento de ARN, específicamente en la maduración del ARN nucleolar pequeño (snoRNA) y el ARN nuclear pequeño(snRNA), y modificación del ARNm de histonas. Semejantes a los cuerpos de Cajal se encuentran los «Geminis de cuerpos enrollados o GEMs (por sus siglas en inglés de Gemini of Coiled Bodies), cuyo nombre se deriva de la constelación de Géminis por su relación casi como de gemelos con los Cuerpos de Cajal. Los GEMs son similares en forma y tamaño a éstos últimos, y de hecho son virtualmente indistinguibles al microscopio. A diferencia de los cuerpos de Cajal, no contienen snRNPs, pero contienen una proteína que se denomina motoneurona superviviente (SMN, por sus siglas en inglés de survivor of motor neurons), cuya función se relaciona con la biogénesis del snRNP. Se cree que los GEMs ayudan a los CBs en la biogénesis del snRNP, aunque también se ha sugerido a partir de evidencias de microscopía que los CBs y los GEMs son diferentes manifestaciones de la misma estructura.
- Dominios PIKA y PTF
Los dominios PIKA, o Asociaciones Cariosómicas Polimórficas de Interface, fueron descritos por primera vez en estudios de microscopía en 1991. Su función era y permanece poco clara, aunque no se piensa que estén asociados con la replicación activa de ADN, transcripción o procesamiento de ARN. Se ha visto que frecuentemente se asocian con dominios discretos definidos por localizaciones densas del factor de transcripción PTF, que promueve la transcripción del ARNnp.
- Cuerpos PML
Los cuerpos PML o de la proteína de la leucemia promielocítica (PML, por sus siglas en inglés de Promyelocytic leukaemia) son cuerpos esféricos que se encuentran dispersos en el nucleoplasma, y que miden alrededor de 0,2-1,0 µm. Se conocen por otros nombres, como dominio «nuclear 10» (ND10), «cuerpos de Kremer», y «dominios oncogénicos PML». A menudo se ven en el núcleo asociados con los cuerpos de Cajal. Se ha sugerido que desempeñan un papel en la regulación de la transcripción.
- Paraspeckles
Descubiertos en 2002, los paraspeckles son compartimentos de forma irregular del espacio intercromatínico del núcleo. Fueron documentados por primera vez en células He La, donde por lo general se encuentran entre 10-30 por núcleo, actualmente se sabe que los paraspeckles también existen en todas las células primarias humanas, los linajes de células transformadas y las secciones de tejidos. Su nombre se deriva de su distribución en el núcleo. El prefijo «para» es una apócope de «paralelo» y «speckles» (mancha o mota, en inglés) se refiere a su proximidad a los «splicing speckles» o motas de ayuste. Los paraspeckles son estructuras dinámicas que se alteran en respuesta a cambios en la actividad celular metabólica. Son dependientes de la transcripción, y en ausencia de transcripción de la ARN Pol II, los paraspeckles desaparecen, y todas las proteínas asociadas que lo componen (PSP1, p54nrb, PSP2, CFI(m)68 y PSF) forman un tapón perinucleolar en forma de cuarto creciente en el nucléolo. Este fenómeno se manifiesta durante el ciclo celular, en el que están presentes en interface y durante toda la mitosis, excepto en telofase. Durante la telofase, cuando los dos núcleos hijos se forman, no hay transcripción por parte de la ARN polimerasa II, de modo que los componentes proteicos forman en su lugar un tapón perinucleolar.
- Speckles
En ocasiones denominados agrupaciones de gránulos intercromatínicos o compartimentos de factores de ayuste, los speckles, manchas o motas, son ricos en ARNnps procedentes del ayuste y otras proteínas del mismo proceso que se necesitan en el procesamiento del pre-ARNm. Debido a los requerimientos variables de la célula, la composición y localización de estos cuerpos cambia de acuerdo a la transcripción de ARNm y a la regulación vía fosforilación de proteínas específicas.
- Cuerpos de escisión
Llamados Cleavage bodies, en inglés, se suelen encontrar asociados a los cuerpos de Cajal, con un diámetro de 0,2 a 1,0 ?m y en número de 1-10 por núcleo. A diferencia de otros cuerpos nucleares, aparecen solamente durante determinados periodos del ciclo celular. Algunos de estos contienen el complejo CPSF-100 (por sus siglas en inglés de cleavage and polyadenylation specificity factor: factor de especificidad para el corte y la poliadenilación), y se pueden observar predominantemente durante las fases S y G, mientras que los que contienen el factor de poliadenilación CstF-64-containing se observan principalmente en la fase S. Están asociados con el clúster de genes de la histona.
- Cuerpos DDX1
Los cuerpos DDX1 son agregados de la proteína DDX1, perteneciente a la familia de helicasas de ARN que contienen el motivo «DEAD box», se encuentran en un número que varía de dos a cuatro. Puesto que parece que estos cuerpos son reclutados en lugares en los que se ha producido daño en el ADN que está hibridando con ADN, parece que estos cuerpos desempeñan un papel en la reparación de zonas con rupturas de doble cadena, facilitando la reparación guiada por patrón de regiones del genoma transcripcionalmente activas.
- Función
La principal función del núcleo celular es controlar la expresión genética y mediar en la replicación del ADN durante el ciclo celular. El núcleo proporciona un emplazamiento para la transcripción en el citoplasma, permitiendo niveles de regulación que no están disponibles en procariotas. Tiene diferentes funciones:
- En el núcleo se guardan los genes en forma de cromosomas (durante la mitosis) o cromatina (durante la interface)
- Organiza los genes en cromosomas lo que permite la división celular
- Transporta los factores de regulación a través de los poros nucleares
- Produce ácido nucleico mensajero (ARNm) que codifica proteínas.
- Produce pre-ribosomas (ARNr) en el nucléolo.
- Compartimentalización celular
La envoltura nuclear permite al núcleo controlar su contenido y separarlo del resto del citoplasma cuando sea necesario. Esto es importante para controlar procesos en cualquiera de los lados de la membrana nuclear. En algunos casos, cuando se precisa restringir un proceso citoplasmático, un participante clave se retira al núcleo, donde interactúa con factores de transcripción para reprimir la producción de ciertas enzimas de la ruta. Este mecanismo regulador tiene lugar en el caso de la glucólisis, una ruta celular en la que se utiliza la glucosa para producir energía. La hexoquinasa es la enzima responsable del primer paso de la glucólisis, produciendo glucosa-6-fosfato a partir de la glucosa. A altas concentraciones de fructosa-6-fosfato, una molécula que se forma posteriormente a partir de la glucosa-6-fosfato, una proteína reguladora retira la hexoquinasa al núcleo, donde forma un complejo con otras proteínas nucleares que reprime la transcripción de los genes implicados en la glucolisis. Para controlar qué genes se deben transcribir, la célula impide el acceso físico de algunos factores de transcripción responsables de regular la expresión génica hasta que son activados por otras rutas de señalización. Esto impide que se den incluso pequeños niveles de expresión génica inadecuada. Por ejemplo, en el caso de los genes controlados por NF-?B, que están implicados en la mayor parte de las respuestas inflamatorias, la transcripción se induce en respuesta a una cascada de señalización celular como la que se inicia con la molécula señalizadora TNF-? uniéndose a un receptor de la membrana celular, lo que produce el reclutamiento de proteínas señalizadoras y finalmente la activación del factor de transcripción NF-?B. Una señal de localización nuclear que posee la proteína NF-?B le permite ser transportada a través del poro nuclear al núcleo, donde estimula la transcripción de los genes diana. La compartimentalización permite a la célula impedir la traducción de ARNm no ayustado. El ARNm contiene intrones que se deben retirar antes de ser traducidos para producir proteínas funcionales. El ayuste se efectúa en el interior del núcleo antes de que el ARNm pueda acceder a los ribosomas para su traducción. Sin el núcleo los ribosomas traducirían ARNm recién transcrito y sin procesar, lo que produciría proteínas mal plegadas y deformadas.
- Expresión génica

- Procesamiento del pre-ARNm
Las moléculas de ARNm recién sintetizadas se conocen como transcritos primarios o pre-ARNm. Posteriormente se deben someter a modificación post-transcripcional en el núcleo antes de ser exportados al citoplasma. El ARNm que aparece en el núcleo sin estas modificaciones acaba degradado en lugar de utilizarse para la traducción en los ribosomas. Las tres modificaciones principales son: La del extremo 5′ (5′ caping), la poliadenilación del extremo 3′ y el ayuste de ARN. Mientras permanece en el núcleo, el pre-ARNm se asocia con varias proteínas en complejos conocidos como ribonucleoproteínas heterogéneas nucleares o hnRNPs. La adición de las modificaciones del extremo 5′ tiene lugar en el momento de la transcripción y es el primer paso en las modificaciones postranscripcionales. La cola de poliadenina 3′ solo se añade una vez que la transcripción está completa. El ayuste (splicing o corte y empalme) de ARN, llevado a cabo por un complejo denominado espliceosoma es el proceso por el que los intrones se retiran del pre-ARNm, permaneciendo únicamente los exones conectados para formar una sola molécula continúa. Este proceso normalmente finaliza tras los dos anteriores, pero puede comenzar antes de que la síntesis esté completa en transcritos con muchos exones. Muchos pre-ARNm’s, incluyendo los que codifican anticuerpos, se pueden cortar y empalmar de múltiples formas para producir diferentes ARNm maduros, que por ello codifican diferentes secuencias de proteínas. Este proceso se conoce como ayuste alternativo, y permite la producción de una gran variedad de proteínas a partir de una cantidad limitada de ADN.
- Dinámica y regulación
- Transporte nuclear

- Ensamblaje y desensamblaje

- Células anucleadas y polinucleadas

- Evolución
Al ser la mejor característica que define la célula eucariota, el origen evolutivo del núcleo ha sido objeto de mucha especulación. Entre las teorías propuestas, se pueden considerar cuatro como las principales, aunque ninguna de ellas ha encontrado un amplio apoyo.
- Teorías endosimbioticas
La teoría conocida como «modelo sintrófico» propone que una relación simbiótica entre arqueas y bacterias creó la primera célula eucariota nucleada. Se establece la hipótesis de que la simbiosis tuvo lugar cuando una arquea antigua similar a los actuales metanógenos fueron invadidos y parasitados por bacterias similares a las actuales myxobacteria, formando eventualmente el núcleo primitivo. Esta teoría es análoga a teoría aceptada del origen de las mitocondrias y cloroplastos eucariotas, de los que se piensa que se han desarrollado por una relación endosimbionte similar entre protoeucariotas y bacterias aerobias. El origen arqueano del núcleo está apoyado por la circunstancia de que tanto arqueas como eucariotas tienen genes similares en ciertas proteínas, incluyendo las histonas. Al observar que las myxobacterias son móviles, pueden formar complejos multicelulares y poseen proteínas G similares a las de eucariotas, también se puede aceptar un origen bacteriano de la célula eucariota. Una propuesta similar establece que una célula similar a la eucariota, el cronocito, apareció en primer lugar, y posteriormente fagocitó arqueas y bacterias para dar lugar al núcleo y a la célula eucariota. Un modelo más controvertido, conocido como eucariogénesis viral afirma que muchos rasgos de la célula eucariota como la presencia de un núcleo que se continúa con la membrana surgieron por la infección de un antepasado procariota por un gran virus de ADN (posiblemente de un Virus nucleocitoplasmáticos de ADN de gran tamaño). Esto está sugerido en base a similitudes entre eucariotas y virus como las hebras lineales de ADN, el procesamiento «caping» del extremo 5′ del ARNm y la fuerte unión a proteínas del ADN (haciendo a las histonas análogas de la envoltura vírica). Una versión de esta propuesta sugiere que el núcleo evolucionó concertadamente con la fagocitosis para dar lugar a un depredador celular primitivo. Otra variante propone que los eucariotas se originaron de arqueas primitivas infectadas por poxvirus, basándose en la similitud de las modernas ADN polimerasas entre éstos y los eucariotas. Se ha sugerido que la cuestión no resuelta de la evolución de la sexualidad pudo estar relacionada con la hipótesis de la eucariogénesis viral.
- Teorías no endosimbioticas
Este modelo propone que las células protoeucariotas evolucionaron a partir de bacterias sin que se diera un estadio simbionte. Este modelo se basa en la existencia de una bacteria moderna perteneciente al filo de las planctomycetes que poseen una estructura nuclear con poros primitivos y otras estructuras compartimentalizadas por membrana. Finalmente, una propuesta muy reciente sugiere que las variantes tradicionales de las teorías endosimbiontes son insuficientes para explicar el origen del núcleo eucariota. Este modelo, denominado la hipótesis de la exomembrana, sugiere que el núcleo se originó en lugar de ello a partir de una célula ancestral original que desarrolló una segunda membrana celular exterior. La membrana interior que encerraba la célula original se convirtió entonces en la membrana nuclear evolucionando para desarrollar estructuras de poro cada vez más elaboradas para el paso de componentes celulares sintetizados internamente, como las subunidades ribosómicas

- ribosomas
Los ribosomas son complejos macromoleculares de proteínas y ácido ribonucleico(ARN) que se encuentran en el citoplasma, en las mitocondrias, en el retículo endoplasmático y en los cloroplastos. Son un complejo molecular encargado de sintetizar proteínas a partir de la información genética que les llega del ADN transcrita en forma de ARN mensajero (ARNm). Sólo son visibles al microscopio electrónico, debido a su reducido tamaño (29 nm en células procariotas y 32 nm en eucariotas). Bajo el microscopio electrónico se observan como estructuras redondeadas, densas a los electrones. Bajo el microscopio óptico se observa que son los responsables de la basofilia que presentan algunas células. Están en todas las células (excepto en los espermatozoides). Los ribosomas están considerados en muchos textos como orgánulos no membranosos, ya que no existen endomembranas en su estructura, aunque otros biólogos no los consideran orgánulos propiamente por esta misma razón. En células eucariotas, los ribosomas se elaboran en el núcleo pero desempeñan su función de síntesis en el citosol. Están formados por ARN ribosómico (ARNr) y por proteínas. Estructuralmente, tienen siempre dos subunidades: la mayor o grande y la menor o pequeña. En las células, estas macromoléculas aparecen en diferentes estados de disociación. Cuando están completas, pueden estar aisladas o formando grupos (polisomas). Las proteínas sintetizadas por los ribosomas actúan principalmente en el citosol; también pueden aparecer asociados al retículo endoplasmático rugoso o a la membrana nuclear, y las proteínas que sintetizan son sobre todo para la secreción. Tanto el ARNr como las subunidades de los ribosomas se suelen nombrar por su coeficiente de sedimentación en unidades Svedberg. En las células eucariotas, los ribosomas del citoplasma alcanzan 80 S. En plastos de eucariotas, así como en procariotas, son 70 S. Los ribosomas mitocondriales son de tamaño variado, entre 55 y 70 S. Subunidad menor del ribosoma. El ARNr es una sola molécula.
- Ribosoma procariota
En la célula procariota, los ribosomas tienen un coeficiente de sedimentación de 70 S. Contienen un 66% de ARNr y se dividen en dos subunidades de distinto tamaño:
- Subunidad mayor: Su coeficiente de sedimentación es 50 S. Tiene dos tipos de ARNr: 5 S (con 120 nucleótidos) y 23 S (2.904 nt), y tiene 31 proteínas como promedio.
- Subunidad menor: Su coeficiente de sedimentación es 30 S. Tiene una sola molécula de ARNr 16 S con 1.542 nucleótidos y contiene 21 proteínas.
- Ribosoma eucariota
En la célula eucariota, los ribosomas tienen un coeficiente de sedimentación de 80 S. Su peso molecular es de 4.194 Kd. Contienen un 60% de ARNr y 40% de proteínas. Al igual que los procariotas se dividen en dos subunidades de distinto tamaño:
- Subunidad mayor: Coeficiente de sedimentación de 60 S. Tres tipos de ARNr: 5 S, 28 S y 5,8 S y tiene 49 proteínas, todas ellas distintas a las de la subunidad menor.
- Subunidad menor: Coeficiente de sedimentación es 40 S. Tiene una sola molécula de ARNr 18 S y contiene 33 proteínas. Dependiendo del organismo eucariota, este ARNr 18 S puede presentar variaciones.
- Ribosoma mitocondrial
Los ribosomas mitocondriales o «mitorribosomas» junto con ARNt y ARNm, son parte del aparato propio de síntesis proteica que tienen las mitocondrias. Son de tamaño variable, desde los 50S de Leishmania hasta 72S en Cándida. Los mitoribosomas de las células animales son 55S y sus dos tipos de ARN ribosómicos, el 12S y 16S, se transcriben a partir de genes del ADN mitocondrial, y son transcritos por una ARN polimerasa mitocondrial específica. Todas las proteínas que forman parte de los ribosomas mitocondriales están codificadas por genes del núcleo celular, que son traducidos en el citosol y transportados hasta las mitocondrias.
- Ribosoma plastidial
Los ribosomas que aparecen en plastos son similares a los procariotas. Son, al igual que los procariotas, 70 S, pero en la subunidad mayor hay un ARNr de 4 S que es equivalente al 5 S procariota. La subunidad mayor 50S tiene unas 33 proteínas y la subunidad menor 30S tiene unas 25 proteínas. La gran mayoría de estas proteínas son homólogas (ortólogas) a las proteínas ribosomales bacterianas y unas pocas son específicas de los cloroplastos.
- Funciones
Los ribosomas son las estructuras supramoleculares encargadas de la síntesis de proteínas, en un proceso conocido como traducción. La información necesaria para esa síntesis se encuentra en el ARN mensajero (ARNm), cuya secuencia de nucleótidos determina la secuencia de aminoácidos de la proteína; a su vez, la secuencia del ARNm proviene de la transcripción de un gen del ADN. El ARN de transferencia lleva los aminoácidos a los ribosomas donde se incorporan al polipéptidos en crecimiento.
- Traducción

- Ribosoma durante la traducción.
Animación de un ribosoma durante el proceso de traducción o síntesis proteica en el retículo endoplasmático El ribosoma lee el ARN mensajero y ensambla los aminoácidos suministrados por los ARN de transferencia a la proteína en crecimiento, proceso conocido como traducción o síntesis de proteínas. Todas las proteínas están formadas por aminoácidos. Entre los seres vivos se han descubierto hasta ahora 20 aminoácidos. En el código genético, cada aminoácido está codificado por uno o varios codones. En total hay 64 codones que codifican 20 aminoácidos y 3 señales de parada de la traducción. Esto hace que el código sea redundante y que haya varios codones diferentes para un mismo aminoácido. La traducción comienza, en general, con el codón AUG que codifica el aminoácido metionina. Al final de la secuencia se ubica un codón que indica el final de la proteína; es el codón de terminación. El código genético es universal porque cada codón codifica el mismo aminoácido para la mayoría de los organismos (no todos). El ribosoma consta de dos partes, la subunidad mayor y una menor, estas salen del núcleo celular por separado. Las subunidades se mantienen unidas por cargas. Al disminuir experimentalmente la concentración de Mg2+, las subunidades tienden a separarse. Por ejemplo, en el citoplasma de una célula eucariota, el proceso con la siguiente secuencia de ARN mensajero sería este:
- AUG le indica que tiene que empezar a ensamblar la proteína. Es un codón de iniciación. Ensambla una metionina.
- GCC es alanina. Toma una alanina y la une.
- AAC es asparagina, lo une con la alanina.
- GGC es glicina, lo ensambla a la asparagina.
- AUG era el símbolo de iniciación, pero el proceso ya ha comenzado. Une una metionina con la glicina anterior.
- CCU es prolina. Ensambla la prolina a la metionina.
- ACU es treonina. Ensambla la treonina con la prolina.
- UAG es terminación. Deja de ensamblar la proteína.
- Por tanto, la cadena polipeptídica ensamblada ha sido: Alanina-Asparagina-Glicina-Metionina-Prolina-Treonina.
Traducción (1) de ARNm por un ribosoma (2) en una cadena polipeptídica (3). El ARNm comienza con un codón de iniciación (AUG) y finaliza con un codón de terminación (UAG).
- retículo endoplásmico
El retículo endoplasmático es un complejo sistema de membranas dispuestas en forma de sacos aplanados y túbulos que están interconectados entre sí compartiendo el mismo espacio interno. Sus membranas se continúan con las de la envuelta nuclear y se pueden extender hasta las proximidades de la membrana plasmática, llegando a representar más de la mitad de las membranas de una célula. Debido a que los ácidos grasos que las componen suelen ser más cortos, son más delgadas que las demás. El retículo organiza sus membranas en regiones o dominios que realizan diferentes funciones. Los dos dominios más fáciles de distinguir son el retículo endoplasmático rugoso, con sus membranas formando túbulos más o menos rectos, a veces cisternas aplanadas, y con numerosos ribosomas asociados, y el retículo endoplasmático liso, sin ribosomas asociados y con membranas organizadas formando túbulos muy curvados e irregulares. La membrana externa de la envuelta nuclear se puede considerar como parte del retículo endoplasmático puesto que es una continuación física de él y se pueden observar ribosomas asociados a ella realizando la traducción. El retículo endoplasmático rugoso y el liso suelen ocupar espacios celulares diferentes como ocurre en los hepatocitos, en las neuronas y en las células que sintetizan esteroides. Sin embargo, en algunas regiones del retículo no existe una segregación clara entre ambos dominios y se aprecian áreas de membrana con ribosomas mezcladas con otras sin ribosomas. La disposición espacial del retículo endoplasmático en las células animales depende de sus interacciones con los microtúbulos, mientras que en las vegetales son los filamentos de actina los responsables. Intervienen en funciones relacionadas con la síntesis proteica, metabolismo de lípidos y algunos esteroides, así como el transporte intracelular. Se encuentra en las células animales y vegetales, pero no en la célula procariota.

- Núcleolo
- Poro nuclear.
- Retículo endoplasmático rugoso (RER).
- Retículo endoplasmático liso (REL).
- Ribosoma en el RE rugoso.
- Proteínas siendo transportadas.
- Vesícula (transporte).
- Aparato de Golgi.
- Lado cis del aparato de Golgi.
- Lado trans del aparato de Golgi.
- Cisternas del aparato de Golgi.
- Clasificación
El retículo endoplasmático rugoso se encuentra unido a la membrana nuclear externa mientras que el retículo endoplasmático liso es una prolongación del retículo endoplasmático rugoso.
- Retículo endoplasmático rugoso
El retículo endoplasmático rugoso tiene esa apariencia debido a los numerososribosomas adheridos a su membrana mediante unas proteínas denominadas «riboforinas». Tiene unos sáculos más redondeados cuyo interior se conoce como «luz del retículo» o «lumen» donde caen las proteínas sintetizadas en él. Está muy desarrollado en las células que por su función deben realizar una activa labor de síntesis, como las células hepáticas o las células delpáncreas.También se le conoce como R.E.R.
- Retículo endoplasmático liso
El retículo endoplasmático liso no tiene ribosomas y participa en el metabolismo de lípidos. También se conoce como R.E.L.
- Funciones
- Biosíntesis proteica: El ARN mensajero proviene de la transcripción del ADN nuclear y es su imagen especular. Al llegar al retículo endoplasmático, se fija a unas estructuras específicas llamadas ribosomas, adheridas al retículo endoplasmático. Allí participa en la síntesis de proteínas, determinando el orden en que se unirán los aminoácidos. información está codificada en forma de tripletes: cada tres bases constituyen un codón que determina un aminoácido. Las reglas de correspondencia entre codones y aminoácidos constituyen el código genético. Los aminoácidos son enviados por el ARN de transferencia, específico para cada uno de ellos, y son trasportados hasta el ARN mensajero, donde se aparean el codón de éste y el anticodón del ARN de transferencia, por complementar de bases, y de ésta forma se sitúan en la posición que les corresponde. Una vez finalizada la síntesis de una proteína, el ARN mensajero queda libre y puede ser leído de nuevo. De hecho, es muy frecuente que antes de que finalice una proteína ya está comenzando otra, con lo cual, una misma molécula de ARN mensajero, está siendo utilizada por varios ribosomas simultáneamente.
- Metabolismo de lípidos: Dado que no tiene ribosomas, en el retículo endoplasmático liso no se sintetizan proteínas. Pero tiene un papel esencial en la síntesis de lípidos de la membrana plasmática, colesterol y derivados de éste, como los ácidos biliares o las hormonas esteroideas.
- Desintoxificación: Es un proceso que se lleva a cabo principalmente en las células del hígado y que consiste en la inactivación de productos tóxicos como drogas, medicamentos o los propios productos del metabolismo celular, por ser liposolubles (hepatocitos).
- Glicosilación: Son reacciones de transferencia de un oligosacárido a las proteínas sintetizadas. Se realiza en la membrana del retículo endoplasmático. De este modo, la proteína sintetizada se transforma en una proteína periférica externa del glucocálix en la reproducción de lisosomas.
- aparato de Golgi
El aparato de Golgi es un orgánulo presente en todas las células eucariotas. Pertenece al sistema de endomembranas. Está formado por unos 80 dictiosomas (dependiendo del tipo de célula), y estos dictiosomas están compuestos por 40 o 60 cisternas (sáculos) aplanadas rodeados de membrana que se encuentran apilados unos encima de otros, y cuya función es completar la fabricación de algunas proteínas. Funciona como una planta empaquetadora, modificando vesículas del retículo endoplasmático rugoso. El material nuevo de las membranas se forma en varias cisternas del aparato de Golgi. Dentro de las funciones que posee el aparato de Golgi se encuentran la glicosilación de proteínas, selección, destinación, glicosilación de lípidos, almacenamiento y distribución de lisosomas, al igual que los peroxisomas, que son vesículas de secreción de sustancias. La síntesis de polisacáridos de la matriz extracelular. Debe su nombre a Camillo Golgi, Premio Nobel de Medicina en 1906 junto a Santiago Ramón y Cajal.

- Núcleo.
- Poro nuclear.
- Retículo endoplasmático rugoso (RER).
- Retículo endoplasmático liso (REL).
- Ribosoma en el RER.
- Proteínas trasportadas.
- Vesícula trasportadora.
- Aparato de Golgi (AG).
- Cisterna del AG.
- Transmembrana de AG.
- Cisterna de AG.
- Vesícula secretora.
- Membrana plasmática.
- Proteína secretada.
- Citoplasma.
- Matriz extracelular.
- Estructura del aparato de Golgi
El aparato de Golgi se compone en estructuras denominadas sáculos. Éstas se agrupan en número variable, habitualmente de 4 a 8, formando el dictiosoma en las plantas. Presentan conexiones tubulares que permiten el paso de sustancias entre las cisternas. Los sáculos son aplanados y curvados, con su cara convexa (externa) orientada hacia el retículo endoplasmático. Normalmente se observan entre 4 y 8, pero se han llegado a observar hasta 60 dictiosomas. Alrededor de la cisterna principal se disponen las vesículas esféricas recién exocitadas. El aparato de Golgi se puede dividir en tres regiones funcionales:
- Región Cis-Golgi: es la más interna y próxima al retículo. De él recibe las vesículas de transición, que son sáculos con proteínas que han sido sintetizadas en la membrana del retículo endoplasmático rugoso (RER), introducidas dentro de sus cavidades y transportadas por el lumen hasta la parte más externa del retículo. Estas vesículas de transición son el vehículo de dichas proteínas que serán transportadas a la cara externa del aparato de Golgi.
- Región medial: es una zona de transición.
- Región Trans-Golgi: es la que se encuentra más cerca de la membrana plasmática. De hecho, sus membranas, ambas unitarias, tienen una composición similar.
Las vesículas provenientes del retículo endoplásmico se fusionan con el cis-Golgi, atravesando todos los dictiosomas hasta el trans-Golgi, donde son empaquetadas y enviadas al lugar que les corresponda. Cada región contiene diferentes enzimas que modifican selectivamente las vesículas según donde estén destinadas. Sin embargo, aún no se han logrado determinar en detalle todas las funciones y estructuras del aparato de Golgi. Diagrama del sistema de endomembranas en una célula eucariota típica.
- Funciones generales
La célula sintetiza un gran número de diversas macromoléculas necesarias para la vida. El aparato de Golgi se encarga de la modificación, distribución y envío de dichas macromoléculas en la célula. Modifica proteínas y lípidos (grasas) que han sido sintetizados previamente tanto en el retículo endoplasmático rugoso como en el liso y los etiqueta para enviarlos a donde corresponda, fuera o dentro de la célula. Las principales funciones del aparato de Golgi vienen a ser las siguientes:
- Modificación de sustancias sintetizadas en el RER: En el aparato de Golgi se transforman las sustancias procedentes del RER. Estas transformaciones pueden ser agregaciones de restos de carbohidratos para conseguir la estructura definitiva o para ser proteolizados y así adquirir su conformación activa. Por ejemplo, en el RER de las células acinosas del páncreas se sintetiza la proinsulina que debido a las transformaciones que sufre en el aparato de Golgi, adquirirá la forma o conformación definitiva de la insulina. Las enzimas que se encuentran en el interior de los dictiosomas son capaces de modificar las macromoléculas mediante glicosilación (adición de carbohidratos) y fosforilación (adición de fosfatos). Para ello, el aparato de Golgi transporta ciertas sustancias como nucleótidos y azúcares al interior del orgánulo desde el citoplasma. Las proteínas también son marcadas con secuencias señal que determinan su destino final, como por ejemplo, la manosa-6-fosfato que se añade a las proteínas destinadas a los lisosomas. Para llevar a cabo el proceso de fosforilación el aparato de Golgi importa moléculas de ATP al interior del lumen, donde las kinasas catalizan la reacción. Algunas de las moléculas fosforiladas en el aparato de Golgi son las apolipoproteínas que dan lugar a las conocidas VLDL que se encuentran en el plasma sanguíneo. Parece ser que la fosforilación de estas moléculas es necesaria para favorecer la secreción de las mismas al torrente sanguíneo.
- Secreción celular: las sustancias atraviesan todos los sáculos del aparato de Golgi y cuando llegan a la cara trans del dictiosoma, en forma de vesículas de secreción, son transportadas a su destino fuera de la célula, atravesando la membrana citoplasmática por exocitosis. Un ejemplo de esto son los proteoglicanos que conforman la matriz extracelular de los animales. El aparato de Golgi es el orgánulo de mayor síntesis de carbohidratos. Esto incluye la producción de glicosaminoglicanos (GAGs), largos polisacáridos que son anclados a las proteínas sintetizadas en el RE para dar lugar a los proteoglicanos. De esto se encargarán las enzimas del Golgi por medio de un residuo de xilosa. Otra forma de marcar una proteína puede ser por medio de la sulfatación de una sulfotransferasa, que gana una molécula de azufre de un donador denominado PAPs. Este proceso tiene lugar en los GAGs de los proteoglicanos así como en los núcleos de las proteínas. Este nivel de sulfatación es muy importante para los proteoglicanos etiquetando funciones y dando una carga neta negativa al proteoglicano.
- Producción de membrana plasmática: los gránulos de secreción cuando se unen a la membrana en la exocitosis pasan a formar parte de esta, aumentando el volumen y la superficie de la célula.
- Formación de los lisosomas primarios.
- Formación del acrosoma de los espermios.
- Vesículas de transporte
Las vesículas formadas en el retículo endoplasmático liso forman, uniéndose entre ellas, agregados tubulo-vesiculares, los cuales son transportados hasta la región cis del aparato de Golgi por proteínas motoras guiadas por microtúbulos donde se fusionan con la membrana de éste, vaciando su contenido en el interior del lumen. Una vez dentro, las moléculas son modificadas, marcadas y dirigidas hacia su destino final. El aparato de Golgi tiende a ser mayor y más numeroso en aquellas células que sintetizan y secretan continuamente sustancias, como pueden ser los linfocitos B y las células secretoras de anticuerpos. Aquellas proteínas destinadas a zonas alejadas del aparato de Golgi son desplazadas hacia la región trans, internándose en una compleja red de membranas y vesículas asociadas denominadas región trans-Golgi. Esta región es donde muchas proteínas son marcadas y enviadas hacia sus correspondientes destinos por medio de alguno de estos 3 tipos diferentes de vesículas, según el marcador que presenten:
- Vesículas de exocitosis
(constitutivas) Este tipo de vesículas contienen proteínas que deben ser liberadas al medio extracelular. Después de internalizarse las proteínas, la vesícula se cierra y se dirige inmediatamente hacia la membrana plasmática, con la que se fusiona, liberando así su contenido al medio extracelular. Este proceso es denominado secreción constitutiva. Los anticuerpos liberados por linfocitos B activados.
- Vesículas de secreción
(reguladas) Este tipo de vesículas contienen también proteínas destinadas a ser liberadas al medio extracelular. Sin embargo, en este caso, la formación de las vesículas va seguida de su almacenamiento en la célula, donde se mantendrán a la espera de su correspondiente señal para activarse. Cuando esto ocurre, se dirigen hacia la membrana plasmática y liberan su contenido como en el caso anterior. Este proceso es secreción regulada. Liberación de neurotransmisores desde las neuronas.
- Vesículas lisosomales
Este tipo de vesículas transportan proteínas destinadas a los lisosomas, unos pequeños orgánulos de degradación en cuyo interior albergan multitud de hidrolasas ácidas, lisosomas de almacenamiento. Estas proteínas pueden ser tanto enzimas digestivas como proteínas de membrana. La vesícula se fusiona con un endosoma tardío y transfiere así su contenido al lisosoma por mecanismos aún desconocidos. Proteasas digestivas destinadas a los lisosomas. >
- Mecanismo de transporte
Microfotografía donde se puede observar el aparato de Golgi como una serie de anillos negros semicirculares apilados cerca de la base. También se pueden observar numerosas vesículas circulares en las proximidades del orgánulo. Los mecanismos de transporte que utilizan las proteínas para trasladarse a través del aparato de Golgi no están muy claros aún, por lo que existen diversas hipótesis para explicar dicho desplazamiento. Actualmente, existen dos modelos predominantes que no son excluyentes entre sí, hasta el punto de ser referidos a veces como el modelo combinado.
- Modelo de maduración de las cisternas: las cisternas del aparato de Golgi llevan cabo un movimiento unidireccional desde la región cis, donde se forman, hasta la región trans, donde son destruidas. Las vesículas del retículo endoplasmático se fusionan con los dictiosomas de la región cis para dar lugar a nuevas cisternas, lo que podría generar el movimiento de las cisternas a través del aparato de Golgi a medida que se van formando nuevas cisternas en la región cis. Este modelo se apoya en el hecho de que se han observado al microscopio estructuras mayores que las vesículas de transporte, tales como las fibras de colágeno, desplazándose a través del aparato de Golgi. Inicialmente, esta hipótesis tuvo una gran acogida y fue la más aceptada hasta los años 80. Los últimos estudios realizados al respecto por la Universidad de Tokio y la Universidad de Chicago con tecnología más avanzada han permitido observar con mayor detalle los compartimentos y el proceso de maduración del aparato de Golgi. Además, existen evidencias de movimientos retrógrados (en dirección cis) de cierto tipo de vesículas (COP1), que transportan proteínas del retículo endoplasmático mediante el reconocimiento de péptidos señales.
- Esquema del transporte en un dictiosoma.
- Vesículas del retículo endoplasmático.
- Vesículas de exocitosis.
- Cisterna.
- Membrana plasmática de la célula.
- Vesícula de secreción.
- Modelo del transporte vesicular: el transporte vesicular asume que el aparato de Golgi es un orgánulo muy estable y estático, dividido en compartimentos que se disponen en dirección cis ? trans. Las vesículas son las encargadas de transportar el material entre el retículo endoplasmático y el aparato de Golgi y entre los diferentes compartimentos de este. Las evidencias experimentales que apoyan esta hipótesis se basan en la gran abundancia de vesículas pequeñas (conocidas técnicamente como vesículas lanzadera) localizadas en las proximidades del aparato de Golgi. La direccionalidad vendría dada por las proteínas trasportadas en el interior de las vesículas, cuyo destino determinaría el movimiento de avance o de retroceso a través del aparato de Golgi, aunque también podría suceder que la direccionalidad no fuera necesaria y el destino de las proteínas viniera ya determinado desde el retículo endoplasmático. Al margen de esto, es probable que el transporte de vesículas se encuentre asociado al citoesqueleto por medio de filamentos de actina, encargados de asegurar la fusión de las vesículas con sus correspondientes compartimentos.
- mitocondrias
Las mitocondrias son orgánulos celulares encargados de suministrar la mayor parte de la energía necesaria para la actividad celular (respiración celular). Actúan, por lo tanto, como centrales energéticas de la célula y sintetizan ATP a expensas de los carburantes metabólicos (glucosa, ácidos grasos y aminoácidos). La mitocondria presenta una membrana exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros llamados porinas o VDAC (canal aniónico dependiente de voltaje), que permiten el paso de moléculas de hasta 10 kDa de masa y un diámetro aproximado de 2 nm Imagen obtenida por microscopía electrónica del tejido pulmonar de un mamífero, se visualizan dos mitocondrias.
- Historia
El descubrimiento de las mitocondrias fue un hecho colectivo. El gran número de términos que se refieren a este orgánulo es prueba de ello: Blefaroplasto, condrioconto, condriómitos, condrioplastos, condriosomas, condriosferas, fila, gránulos fucsinofílicos, Korner, Fadenkörper, mitogel, cuerpos parabasales, vermículas, sarcosomas, cuerpos intersticiales, plasmosomas, plastocondrios, bioblastos. Cowdry intentó en 1918, en un trabajo luego citado por Lehninger, sistematizar y unificar todos los términos. Probablemente las primeras observaciones se deben al botánico suizo Kolliker, quien en 1880-1888 anotó la presencia de unos gránulos en células musculares de insectos a los que denominó sarcosomas. Llegó incluso a la conclusión de que presentaban membrana. En 1882, el alemán Walther Flemming descubrió una serie de inclusiones a las que denominófila. En 1884 también fueron observados por Richard Altmann, quien más tarde en su obra publicada en Leipzig Die Elementarorganismen describe una serie de corpúsculos que observa mediante una tinción especial que incluye fucsina. Especula que se trata de una suerte de parásitos independientes, con su propio metabolismo y los denomina bioblastos. El hallazgo fue rechazado como un artefacto de la preparación, y sólo más tarde fue reconocido como mitocondrias por N.H. Cowdry (1916). También los «plastídulos» del protozoólogo italiano Leopoldo Maggi podrían tratarse de observaciones tempranas de mitocondrias. Sin embargo, el nombre de mitocondria, que es el que alcanzó mayor fortuna, se debe a Carl Benda, quien en 1889denominó así a unos gránulos que aparecían con gran brillo en tinciones de cristal violeta y alizarina, y que anteriormente habían sido denominados «citomicrosomas» por Velette St. George. En 1904 F. Meves confirma su presencia en una planta, concretamente en células del tapete de la antera de Nymphaea, y en 1913 Otto Heinrich Warburg descubre la asociación con enzimas de la cadena respiratoria, aunque ya Kingsbury, en 1912 había relacionado estos orgánulos con la respiración celular. En 1934 fueron aisladas por primera vez a partir de homogeneizados de hígado y en 1948 Hogeboon, Schneider y Palade establecen definitivamente la mitocondria como el lugar donde se produce la respiración celular. La presencia del ADN mitocondrial fue descubierta por Margit M. K. Nass y Sylvan Nass en 1963.
- Estructura y composición

- Estructura de una mitocondria
La morfología de la mitocondria es difícil de describir puesto que son estructuras muy plásticas que se deforman, se dividen y fusionan. Normalmente se las representa en forma alargada. Su tamaño oscila entre 0,5 y 1 ?m de diámetro y hasta 7 ?m de longitud. Su número depende de las necesidades energéticas de la célula. Al conjunto de las mitocondrias de la célula se le denomina condrioma celular. Las mitocondrias están rodeadas de dos membranas claramente diferentes en sus funciones y actividades enzimáticas, que separan tres espacios: el citosol, el espacio intermembrana y la matriz mitocondrial.
- Membrana externa
Es una bicapa lipídica exterior permeable a iones, metabolitos y muchos polipéptidos. Eso es debido a que contiene proteínas que forman poros, llamadas porinas o VDAC (de canal aniónico dependiente de voltaje), que permiten el paso de grandes moléculas de hasta 5.000 dalton y un diámetro aproximado de 20 Å. La membrana externa realiza relativamente pocas funciones enzimáticas o de transporte. Contiene entre un 60 y un 70% de proteínas.
- Membrana interna
La membrana interna contiene más proteínas, carece de poros y es altamente selectiva; contiene muchos complejos enzimáticos y sistemas de transporte transmembrana, que están implicados en la translocación de moléculas. Esta membrana forma invaginaciones o pliegues llamados crestas mitocondriales, que aumentan mucho la superficie para el asentamiento de dichas enzimas. En la mayoría de los eucariontes, las crestas forman tabiques aplanados perpendiculares al eje de la mitocondria, pero en algunos protistas tienen forma tubular o discoidal. En la composición de la membrana interna hay una gran abundancia de proteínas (un 80%), que son además exclusivas de este orgánulo:
- La cadena de transporte de electrones, compuesta por cuatro complejos enzimáticos fijos y dos transportadores de electrones móviles:
- Complejo I o NADH deshidrogenasa que contiene flavina mononucleótido (FMN).
- Complejo II o succinato deshidrogenasa; ambos ceden electrones al coenzima Q o ubiquinona.
- Complejo III o citocromo bc1 que cede electrones al citocromo c.
- Complejo IV o citocromo c oxidasa que cede electrones al O2 para producir dos moléculas de agua.
- Un complejo enzimático, el canal de H+ ATP sintasa que cataliza la síntesis de ATP (fosforilación oxidativa).
- Proteínas transportadoras que permiten el paso de iones y moléculas a su través, como ácidos grasos, ácido pirúvico, ADP, ATP, O2 y agua; pueden destacarse:
- Nucleótido de adenina translocasa. Se encarga de transportar a la matriz mitocondrial el ADP citosólico formado durante las reacciones que consumen energía y, paralelamente transloca hacia el citosol el ATP recién sintetizado durante la fosforilación oxidativa.
- Fosfato translocasa. Transloca fosfato citosólico junto con un hidrón a la matriz; el fosfato es esencial para fosforilar el ADP durante la fosforilación oxidativa.
- Espacio intermembranoso
Entre ambas membranas queda delimitado un espacio intermembranoso que está compuesto de un líquido similar al hialoplasma; tienen una alta concentración de protones como resultado del bombeo de los mismos por los complejos enzimáticos de la cadena respiratoria. En él se localizan diversas enzimas que intervienen en la transferencia del enlace de alta energía del ATP, como la adenilato kinasa o la creatina quinasa. También se localiza la carnitina, una molécula implicada en el transporte de ácidos grasos desde el citosol hasta la matriz mitocondrial, donde serán oxidados (beta-oxidación).
- Matriz mitocondrial
- La matriz mitocondrial o mitosol contiene menos moléculas que el citosol, aunque contiene iones, metabolitos a oxidar, ADN circular bicatenario muy parecido al de las bacterias, ribosomas tipo 55S (70S en vegetales), llamados mitorribosomas, que realizan la síntesis de algunas proteínas mitocondriales, y contiene ARN mitocondrial; es decir, tienen los orgánulos que tendría una célula procariota de vida libre. En la matriz mitocondrial tienen lugar diversas rutas metabólicas clave para la vida, como el ciclo de Krebs y la beta-oxidación de los ácidos grasos; también se oxidan los aminoácidos y se localizan algunas reacciones de la síntesis de urea y grupos hemo.
- Función
La principal función de las mitocondrias es la oxidación de metabolitos (ciclo de Krebs, beta-oxidación de ácidos grasos) y la obtención de ATP mediante la fosforilación oxidativa, que es dependiente de la cadena transportadora de electrones; el ATP producido en la mitocondria supone un porcentaje muy alto del ATP sintetizado por la célula. También sirve de almacén de sustancias como iones, agua y algunas partículas como restos de virus y proteínas.
- Origen
La científica estadounidense Lynn Margulis, junto con otros científicos, recuperó en torno a 1980 una antigua hipótesis, reformulándola como teoría endosimbioticas. Según esta versión actualizada, hace unos 1.500 millones de años, una célula procariota capaz de obtener energía de los nutrientes orgánicos empleando el oxígeno molecular como oxidante, se fusionó en un momento de la evolución con otra célula procariota o eucariota primitiva al ser fagocitada sin ser inmediatamente digerida, un fenómeno frecuentemente observado. De esta manera se produjo una simbiosis permanente entre ambos tipos de seres: la procariota fagocitada proporcionaba energía, especialmente en forma de ATP y la célula hospedadora ofrecía un medio estable y rico en nutrientes a la otra. Este mutuo beneficio hizo que la célula invasora llegara a formar parte del organismo mayor, acabando por convertirse en parte de ella: la mitocondria. Otro factor que apoya esta teoría es que las bacterias y las mitocondrias tienen mucho en común, tales como el tamaño, la estructura, componentes de su membrana y la forma en que producen energía, etc. Esta hipótesis tiene entre sus fundamentos la evidencia de que las mitocondrias poseen su propio ADN y están recubiertas por su propia membrana. Otra evidencia que sostiene esta hipótesis es que el código genético del ADN mitocondrial no suele ser el mismo que el código genético del ADN nuclear. A lo largo de la historia común la mayor parte de los genes mitocondriales han sido transferidos al núcleo, de tal manera que la mitocondria no es viable fuera de la célula hospedadora y ésta no suele serlo sin mitocondrias.
- Enfermedades mitocondriales
El ADN mitocondrial humano contiene información genética para 13 proteínas mitocondriales y algunos ARN; no obstante, la mayoría de las proteínas de las mitocondrias proceden de genes localizados en el ADN del núcleo celular y que son sintetizadas por ribosomas libres del citosol y luego importadas por el organelo. Se han descrito más de 150 enfermedades mitocondriales, como la enfermedad de Luft o la neuropatía óptica hereditaria de Leber. Tanto las mutaciones del ADN mitocondrial, como del ADN nuclear dan lugar a enfermedades genéticas mitocondriales, que originan un mal funcionamiento de procesos que se desarrollan en las mitocondrias, como alteraciones de enzimas, ARN, componentes de la cadena de transporte de electrones y sistemas de transporte de la membrana interna; muchas de ellas afectan al músculo esquelético y al sistema nervioso central. El ADN mitocondrial puede dañarse con los radicales libres formados en la mitocondria; así, enfermedades degenerativas relacionadas con el envejecimiento, como la enfermedad de Parkinson, la enfermedad de Alzheimer y las cardiopatías pueden tener relaciones con lesiones mitocondriales.
- Pérdida de mitocondrias por evolución
Existen protistas sin mitocondrias que carecen de ellas por una pérdida secundaria o una degeneración de las mismas, para adaptarse a un modo de vida parásito, intracelular o anaerobio.
- cloroplastos
Los cloroplastos son los orgánulos celulares que en los organismos eucariontes fotosintetizadores se ocupan de la fotosíntesis. Están limitados por una envoltura formada por dos membranas concéntricas y contienen vesículas, los tilacoides, donde se encuentran organizados los pigmentos y demás moléculas que convierten la energía lumínica en energía química, como la clorofila. El término cloroplastos sirve alternativamente para designar a cualquier plasto dedicado a la fotosíntesis, o específicamente a los plastos verdes propios de las algas verdes y las plantas.

- Estructura de un cloroplasto
Células vegetales en las que son visibles los cloroplastos.
- Estructura
El cloroplasto está rodeado de dos membranas, que poseen una diversa estructura continua que delimita completamente el cloroplasto. Ambas se separan por un espacio intermembranoso llamado a veces indebidamente espacio periplastidial. La membrana externa es muy permeable gracias a la presencia de porinas, pero en menor medida que la membrana interna, que contiene proteínas específicas para el transporte. La cavidad interna llamada estroma, en la que se llevan a cabo reacciones de fijación de CO2, contiene ADN circular, ribosomas (de tipo 70S, como los bacterianos), gránulos de almidón, lípidos y otras sustancias. También, hay una serie de sáculos delimitados por una membrana llamados tilacoides, que en los cloroplastos de las plantas terrestres se organizan en apilamientos llamados grana (plural de granum, grano). Las membranas de los tilacoides contienen sustancias como los pigmentos fotosintéticos (clorofila, carotenoides, xantófilas) y distintos lípidos; proteínas de la cadena de transporte de electrones fotosintética y enzimas, como la ATP-sintetasa. Al observar la estructura del cloroplasto y compararlo con el de la mitocondria, se nota que ésta tiene dos sistemas de membrana, delimitando un compartimento interno (matriz) y otro externo, el espacio perimitocondrial; por su parte, el cloroplasto tiene tres, que forman tres compartimentos, el espacio intermembrana, el estroma y el espacio intratilacoidal.
- Plasto glóbulos
Como parte de la estructura del cloroplasto, también se pueden encontrar plasto glóbulos, que se desprenden de los tilacoides y están rodeados de una membrana similar a la de los tilacoides, y en su interior son gotas compuestas por moléculas orgánicas entre las que preponderan ciertos lípidos. La función de las moléculas del plasto glóbulos todavía se está estudiando.
- Funciones
Cloroplasto obtenido mediante microscopía electrónica. El cloroplasto es el orgánulo donde se realiza la fotosíntesis de los organismos eucariotas autótrofos. El conjunto de reacciones de la fotosíntesis es realizada gracias a todo un complejo de moléculas presentes en el cloroplasto, una en particular, presente en la membrana de los tilacoides, es la responsable de tomar la energía del Sol, es llamada clorofila a. Existen dos fases, que se desarrollan en compartimentos distintos:
- Fase luminosa: Se realiza en la membrana de los tilacoides, donde se halla la cadena de transporte de electrones y la ATP-sintetasa responsables de la conversión de la energía lumínica en energía química (ATP) y de la generación poder reductor (NADPH).
- Fase oscura: Se produce en el estroma, donde se halla el enzima RuBisCO, responsable de la fijación del CO2mediante el ciclo de Calvin.
- Pigmentos
Un cromóforo es un material que absorbe la luz de ciertos colores, reflejando la luz de otros. La luz absorbida por los cromóforos de la membrana tilacoide de los cloroplastos es utilizada como fuente de energía que impulsa la fotosíntesis. La clorofila a es un cromóforo presente en todos los cloroplastos (y en las cianobacterias de las que se originaron). Las moléculas capaces de absorber luz de algunos colores y reflejar luz de otros se llaman cromóforos, en plantas, los cromóforos están unidos a otras moléculas (proteínas) que les modifican un poco el color de luz absorbido, al complejo formado por cromóforo + proteína se lo llama pigmento, a los fines de este texto trataremos a los cromóforos con el nombre de «pigmentos» ). La clorofila a absorbe luz de colores rojo y azul, reflejando principalmente el verde (de la luz visible). Pero no es el único pigmento, en la membrana de los tilacoides se encuentran diferentes pigmentos que absorben luz de algunos colores con el fin último de impulsar la fotosíntesis. De aquéllos, los que no son clorofila a se llaman pigmentos accesorios. Los pigmentos accesorios permiten captar la energía de la luz de colores diferentes de los captados por la clorofila a. Por ejemplo, se han presentado pequeñas variaciones en la estructura química de la clorofila a debidas a la evolución, estas variaciones son pigmentos accesorios llamados clorofila b, clorofila c1, etc., y captan luz de colores ligeramente diferentes de los que capta la clorofila a, reflejando siempre, principalmente, en la gama del verde. Las demás clorofilas no se encuentran en todos los eucariotas fotosintéticos sino en algunos grupos cuyo cloroplasto desciende de un ancestro común, y comparten casi la vía biosintética con la clorofila a, con un pequeño cambio en la vía que da una clorofila diferente. Hay otros pigmentos accesorios, que no necesariamente se sintetizan por las mismas vías que las clorofilas y por lo tanto su estructura química no es similar a la de ellas, que absorben luz de otros colores, y pueden presentar también sus variaciones debidas a la evolución. Son pigmentos accesorios muy comunes, por ejemplo, los diferentes carotenoides (que captan luz de las gamas verde-azuladas, y reflejan la luz roja, naranja y amarilla). En la membranade los tilacoides, en cada complejo que realiza fotosíntesis sólo un par de moléculas de clorofila a (un dímero) son las responsables de impulsar el proceso de fotosíntesis, el resto de la clorofila a y de los pigmentos accesorios se encuentra alrededor de ese par formando «complejos antena» que captan, de la luz que les llega, los colores que les están permitidos, y le transfieren esa energía al par central. Luego transcurre la fotosíntesis por la fase lumínica y luego la fase oscura. Cada pigmento le da un color diferente a la planta, y a veces llegan a enmascarar el color verde que refleja la clorofila a, siempre presente. Por ejemplo las «algas verdes» tienen principalmente clorofilas, mientras que las algas pardas tienen además fucoxantina que les da su color característico. Debido a que hay hábitats donde la intensidad de luz es muy baja en los colores que capta la clorofila a y más alta en otros colores, los pigmentos accesorios permiten que la planta explore hábitats que de otra forma serían difíciles de alcanzar: así por ejemplo, como la luz azul es la que tiene la mayor penetración en el agua, las algas rojas, que contienen varios pigmentos que absorben los colores azulados (y reflejan los rojos), pueden permitirse vivir en el mar a mayores profundidades que las demás algas. En el mar, la concentración de pigmentos fotosintéticos (en particular de clorofila a) está relacionada con la densidad de algas, por lo que su estimación es muy utilizada para estimar la densidad de algas en relación a la profundidad y al área, y se utilizan técnicas de sensores satelitales (que pueden reconocer los colores absorbidos por los pigmentos) para este propósito.
- Alga verde. Su color es dado principalmente por las clorofilas que poseen en los tilacoides de sus cloroplastos.
- Alga parda. Su color es dado principalmente por el pigmento accesorio llamado fucoxantina, presente en sus cloroplastos.
- Alga roja. Su color es dado por varios pigmentos accesorios que captan principalmente los colores azulados.
- En animales
Elysia chlorotica se ve de color verde luego de haber adquirido la capacidad de realizar fotosíntesis. Hay animales que pueden adquirir cloroplastos por un proceso diferente de la endosimbiosis y que no se heredan. Por un proceso llamado cleptoplastia los organismos heterótrofos consumen y retienen los cloroplastos de un organismo fotosintético. Por ejemplo en el «caracol de mar» sarcoglosso Elysia chlorotica, que es el organismo donde más se estudió este suceso, los cloroplastos se consumen junto con las algas que forman parte del alimento del organismo, el resto del alga se degrada y los cloroplastos se secuestran (se mantienen dentro del citoplasma de las células que debían degradarlos, pasando a ser «cleptoplastos»), de esta forma forman parte de los tejidos del organismo que gana la habilidad de realizar fotosíntesis por un tiempo que puede llegar a ser de varios meses. La eficiencia de la fotosíntesis de estos cleptoplastos es tan alta que si la intensidad de luz es buena, estos moluscos no necesitan alimentarse. Las bases de la longevidad del cleptoplasto y la forma en que son integrados al metabolismo del hospedador son áreas de intensa investigación.
- Otros tipos de plastos
Tipos de plástidos, o plastos.
- Plasto
- Proplasto
- Etioplasto
- Cromoplasto
- Leucoplasto
- Amiloplasto
- Estatolito
- Oleoplasto
- Apicoplasto
- Gerontoplasto
- Origen
Los cloroplastos se originan por un proceso denominado simbiogénesis, en donde se produce la unión quimérica entre un huésped protista heterótrofo biflagelado, probablemente fagótrofo, y una bacteria fotosintética oxigénica endosimbionte, esto significa que el primer plasto desciende directamente de una cianobacteria. Esto pudo ser un evento único en la historia de la vida y daría un respaldo a la monofilia del clado Primoplantae (primera planta) o Archaeplastida (el antiguo plasto), además equivale al origen de la primera célula vegetal, cuyos cloroplastos son los ancestros de todos los plastos existentes, incluyendo aquellos de otros grupos como los cromistas, dinoflagelados y alveolados. La filogenia de las cianobacterias aún no está consensuada. Una versión sobre las relaciones filogenéticas en base a secuencias moleculares es la siguiente (grupos en comillas son parafiléticos):

- Evolución y filogenia
La aparición de los cloroplastos parece ser un evento único, de tal manera que todos los tipos de plastos actuales, tanto de plantas como de todas las algas, descienden en última instancia del este primero cloroplasto (Archaeplastida) en un proceso denominado endosimbiosis primaria. Sin embargo, los plastos tienen una compleja historia evolutiva, con múltiples eventos endosimbióticos, originándose grupos de algas por endosimbiosis secundaria a partir de la simbiogénesis entre un protista biflagelado con un alga clorofita o rodofita, y eventos de endosimbiosis terciaria en varios dinoflagelados. No hay consenso sobre el número de eventos endosimbióticos, ni las exactas relaciones filogenéticas entre todos los eucariontes fotosintéticos; pero en líneas generales las principales líneas evolutivas son las siguientes:

- lisosomas
Los lisosomas son orgánulos relativamente grandes, formados por el retículo endoplasmático rugoso y luego empaquetadas por el complejo de Golgi, que contienen enzimas hidrolíticas y proteolíticas que sirven para digerir los materiales de origen externo (heterofagia) o interno (autofagia) que llegan a ellos. Es decir, se encargan de la digestión celular. Son estructuras esféricas rodeadas de membrana simple. Son bolsas de enzimas que si se liberasen, destruirían toda la célula. Esto implica que la membrana lisosómica debe estar protegida de estas enzimas. El tamaño de un lisosoma varía entre 0.1-1.2 ?m. En un principio se pensó que los lisosomas serían iguales en todas las células, pero se descubrió que tanto sus dimensiones como su contenido son muy variables. Se encuentran en todas las células animales. No se ha demostrado su existencia en células vegetales.
- Las enzimas lisosomales
El pH en el interior de los lisosomas es de 4,8 (bastante menor que el del citosol, que es neutro) debido a que las enzimas proteolíticas funcionan mejor con un pH ácido. La membrana del lisosoma estabiliza el pH bajo bombeando iones (H+) desde el citosol, así mismo, protege al citosol e igualmente al resto de la célula de las enzimas digestivas que hay en el interior del lisosoma. Las enzimas lisosomales son capaces de digerir bacterias y otras sustancias que entran en la célula por fagocitosis, u otros procesos de endocitosis. Los lisosomas utilizan sus enzimas para reciclar los diferentes orgánulos de la célula, englobándolos, digiriéndolos y liberando sus residuos en el citosol. De esta forma los orgánulos de la célula se están continuamente reponiendo. El proceso de digestión de los orgánulos se llama autofagia. Por ejemplo, las células hepáticas se reconstituyen por completo una vez cada dos semanas. Las enzimas más importantes del lisosoma son:
- Lipasas, que digiere lípidos;
- Glucosidasa, que digiere carbohidratos;
- Proteasas, que digiere proteínas;
- Nucleasas, que digiere ácidos nucleicos.
- Formación de lisosomas primarios

Los lisosomas primarios son orgánulos derivados del sistema de endomembranas. Cada lisosoma primario es una vesícula que brota del aparato de Golgi, con un contenido de enzimas hidrolíticas (hidrolasas). Las hidrolasas son sintetizadas en el retículo endoplasmático rugoso y viajan hasta el aparato de Golgi por transporte vesicular. Allí sufren una glicosilación terminal (proceso químico en el que se adiciona un carbohidrato a otra molécula) de la cual resultan con cadenas glucídicas ricas en manosa-6-fosfato (manosa-6-P). La manosa-6-P es el marcador molecular, la «estampilla» que dirige a las enzimas hacia la ruta de los lisosomas. Se ha estudiado una enfermedad en la cual las hidrolasas no llevan su marcador; las membranas del aparato de Golgi no las reconocen como tales y las empaquetan en vesículas de secreción para ser exocitadas. Quienes padecen esta enfermedad acumulan hidrolasas en el medio extracelular, mientras sus células carecen de ellas. Lisosomas secundarios y digestión celular Los lisosomas primarios contienen una variedad de enzimas hidrolíticas capaces de degradar casi todas las moléculas orgánicas. Estas hidrolasas se ponen en contacto con sus sustratos cuando los lisosomas primarios se fusionan con otras vesículas. El producto de la fusión es un lisosoma secundario. Por lo tanto, la digestión de moléculas orgánicas se lleva a cabo en los lisosomas secundarios, ya que éstos contienen a la vez los sustratos y las enzimas capaces de degradarlos. Existen diversas formas de lisosomas secundarios, según el origen de la vesícula que se fusiona con el lisosoma primario:
- Fagolisosomas. Se originan de la fusión del lisosoma primario con una vesícula procedente de la fagocitosis, denominada fagosoma. Se encuentran, por ejemplo, en los glóbulos blancos, capaces de fagocitar partículas extrañas que luego son digeridas por estas células.
- Endosomas tardíos. Surgen al unirse los lisosomas primarios con materiales provenientes de los endosomas tempranos. Los endosomas tempranos contienen macromoléculas que ingresan por los mecanismos de endocitosis inespecífica y endocitosis mediada por receptor. Este último es utilizado por las células para incorporar, por ejemplo, las lipoproteínas de baja densidad o LDL.
- Autofagolisosomas. Es el producto de la fusión entre un lisosoma primario y una vesícula autofágica o autofagosoma. Algunos orgánulos citoplasmáticos son englobados en vesículas, con membranas que provienen de las cisternas del retículo endoplasmático, para luego ser reciclados cuando estas vesículas autofágicas se unen con los lisosomas primarios.
Lo que queda del lisosoma secundario después de la absorción es un cuerpo residual. Los cuerpos residuales contienen desechos no digeribles que en algunos casos se exocitan y en otros no, acumulándose en el citosol a medida que la célula envejece. Un ejemplo de cuerpos residuales son los gránulos de lipofuscina que se observan en células de larga vida, como las neuronas.
- Enfermedades lisosómicas
Son enfermedades causadas por la disfunción de alguna enzima lisosómica o por la liberación incontrolada de dichas enzimas en el citosol, lo que produce la lisis de la célula. En algunos casos, la liberación de las enzimas cumple un papel fisiológico, permitiendo la reabsorción de estructuras que ya no son útiles, por ejemplo la cola de los renacuajos durante la metamorfosis.
- Enfermedades de almacenamiento lisosómico
En las enfermedades de almacenamiento lisosómico, alguna enzima del lisosoma tiene actividad reducida o nula debido a un error genético y el substrato de dicho enzima se acumula y deposita dentro del lisosoma que aumentan de tamaño a causa del material sin digerir, lo cual interfiere con los procesos celulares normales; algunas de estas enfermedades son:
- Esfingolipidosis. Son enfermedades causadas por la disfunción de algunas de las enzimas de la ruta de degradación de los esfingolípidos. Dado que los esfingolípidos abundan en el cerebro, varias de estas enfermedades cursan con retraso mental severo y muerte prematura; entre ellas hay que destacar la enfermedad de Tay-Sachs, la enfermedad de Gaucher, la enfermedad de Niemann-Pick, la enfermedad de Krabbe, la fucosidosis, etc.
- Carencia de lipasa ácida. La lipasa ácida es una enzima fundamental en el metabolismo de los triglicéridos y del colesterol, que se acumulan en los tejidos. La disfunción de esta enzima provoca dos enfermedades, la enfermedad de almacenamiento de ésteres de colesterol, en que la enzima presenta muy poca actividad, y la enfermedad de Wolman, en que la enzima es totalmente inactiva.
- Glucogenosis tipo II o enfermedad de Pompe. Es un defecto de la ? (1-4) glucosidasa ácida lisosómica, también denominada maltasa ácida. El glucógeno aparece almacenado en lisosomas. En niños destaca por producir insuficiencia cardíaca al acumularse en el músculo cardíaco causando cardiomegalia. En adultos el acúmulo es más acusado en músculo esquelético.
- Mucopolisacaridosis. Causadas por la ausencia o el mal funcionamiento de las enzimas necesarias para la degradación moléculas llamadas glicosoaminoglicanos o glucosaminglucanos (antes llamadas mucopolisacáridos). Destacan la mucopolisacaridosis tipo I, también conocida como gargolismo o enfermedad de Hurler, en la que existe un defecto de la enzima ?-1-iduronidasa, y la mucopolisacaridosis de tipo II o síndrome de Hunter, causada por un error en la enzimaiduronato-2-sulfatasa. El -síndrome Sanfilippo– MPS III, está relacionado con la acumulación de N-heparan Sulfatasa.
- peroxisomas
Los peroxisomas son orgánulos citoplasmáticos muy comunes en forma de vesículas que contienen oxidasas y catalasas. Estas enzimas cumplen funciones de detoxificación celular. Como la mayoría de los orgánulos, los peroxisomas solo se encuentran en células eucariotas. Fueron descubiertos en 1965 por Christian de Duve y sus colaboradores. Inicialmente recibieron el nombre de microcuerpos y están presentes en todas las células eucariotas.
- Función
Los peroxisomas tienen un papel esencial en el metabolismo lipídico, en especial en el acortamiento de los ácidos grasos de cadena muy larga, para su completa oxidación en las mitocondrias, y en la oxidación de la cadena lateral del colesterol, necesaria para la síntesis de ácidos biliares; también interviene en la síntesis de ésteres lipídicos del glicerol (fosfolípidos y triglicéridos) e isoprenoides; también contienen enzimas que oxidan aminoácidos, ácido úrico y otros sustratos utilizando oxígeno molecular con formación de agua oxigenada:
| RH2 + O2 ? R + H2O2 |
|---|
El agua oxigenada es un producto tóxico, que se degrada rápidamente dentro del propio peroxisoma por la enzima oxidativa catalasa en agua y oxígeno usando como intermediarios de ciertas sustancias orgánicas (en la ecuación la variable R’). lo cual ayuda al buen funcionamiento de la célula
| H2O2 + R’H2 ? R’ + 2H2O |
|---|
La catalasa es también capaz de utilizar el peróxido de hidrógeno para reacciones de oxidación, como por ejemplo, la oxidación de sustancias tóxicas como los fenoles, etanol, formaldehído, entre otros, las cuales son posteriormente eliminadas. Tal es el mecanismo de detoxificación realizada por el hígado y los riñones, por ejemplo. En las plantas son el asiento de una serie de reacciones conocidas como fotorrespiración.
- Sentido evolutivo
Actualmente los científicos postulan que los peroxisomas han prevalecido desde su aparición como adaptación contra los continuos efectos tóxicos a los que se expone una célula que mantiene un metabolismo aerobio y produce accidentalmente especies reactivas del oxígeno (ROS). Estas especies químicas reaccionan rápidamente con elementos fundamentales para la estabilidad celular como el ADN, de ahí que se les atribuya un papel crítico en el envejecimiento y la pérdida del control del ciclo celular que puede desembocar en tumores y cáncer. En particular, las ROS puede desequilibrar el estado de reducción del citoplasma, lo que provocaría un bloqueo de la cadena transportadora de electrones mitocondrial y la parada transitoria en la producción de energía. En plantas, especialmente en las de metabolismo C3, los peroxisomas son relevos importantes en la ruta bioquímica conocida como fotorrespiración. La función que cumple esta ruta resulta paradójica, ya que parece haber evolucionado para aliviar los efectos perjudiciales de la actividad oxigenasa de la ribulosa-1,5-bisfosfato carboxilasa oxigenasa o RuBisCO (la principal enzima asimiladora de carbono inorgánico en las plantas) sin que en ésta hayan ocurrido aparentemente modificaciones ventajosas dirigidas a un incremento del rendimiento en la actividad carboxilasa. Ya que la fotorrespiración implica no sólo una recuperación de carbono sino también un consumo de energía, algunos autores piensan que la fotorrespiración pudiera resultar ventajosa en situaciones ambientales (altas temperaturas e intensidad luminosa) que requieran un desacoplamiento en la producción de energía como medio para subsanar la producción masiva de radicales libres, esta vez productos de la actividad forzada del aparato fotosintético situado en los cloroplastos. De este modo quedaría salvaguardada la hipótesis inicial en la que proponían el papel accesorio del peroxisoma en la destrucción de las ROS.

- Enfermedades peroxisomales
Se conocen más de 25 enfermedades relacionadas con la disfunción de las actividades enzimáticas de los peroxisomas, conocidas como anomalías de la biogénesis de peroxisomas (PBD). También tenemos la enfermedad de adrenoleucodistrofia ligado al cromosoma X Se trata de enfermedades hereditarias autosómicas recesivas poco frecuentes caracterizadas por alteraciones en el cerebro, riñones, hígado y esqueleto. La más grave es la enfermedad de Zellweger, debida a la ausencia de peroxisomas funcionales, ya que fallan los mecanismos de importación de los enzimas al interior del peroxisoma. Otras enfermedades son causadas por el error de un solo enzima o por defectos en los componentes del transporte de la membrana peroxisomal
- vacuolas
Una vacuola es un orgánulo celular presente en todas las células de plantas y hongos. También aparece en algunas células protistas y de otras eucariotas. Las vacuolas son compartimentos cerrados o limitados por la membrana plasmática ya que contienen diferentes fluidos, como agua o enzimas, aunque en algunos casos puede contener sólidos. La mayoría de las vacuolas se forman por la fusión de múltiples vesículas membranosas. El orgánulo no posee una forma definida, su estructura varía según las necesidades de la célula en particular. Las vacuolas que se encuentran en las células vegetales son regiones rodeadas de una membrana (tonoplasto o membrana vacuolar) y llenas de un líquido muy particular llamado jugo celular. La célula vegetal inmadura contiene una gran cantidad de vacuolas pequeñas que aumentan de tamaño y se van fusionando en una sola y grande, a medida en que la célula va creciendo. En la célula madura, el 90 % de su volumen puede estar ocupado por una vacuola, con el citoplasma reducido a una capa muy estrecha apretada contra la pared celular.
- Origen de las vacuolas vegetales
Desde hace mucho tiempo se ha considerado que las vacuolas se forman del retículo endoplasmático. Cuando se evidenció que eran muy parecidas a los lisosomas de las células animales se llegó a la conclusión, de que las vacuolas de por lo menos algunas células vegetales tenían un origen similar al de los lisosomas animales. La formación de los lisosomas está asociada una región del citoplasma muy especializada llamada GERL, formado por el complejo de Golgi, el retículo endoplasmático y los lisosomas. Esta asociación de membranas se ha encontrado también en algunas células vegetales, por lo que el origen de las vacuolas podría ser el mismo que el de los lisosomas animales.

- Contenido vacuolar
En el interior de las vacuolas, en el jugo celular, se encuentran una gran cantidad de sustancias. La principal de ellas es el agua, junto a otros componentes que varían según el tipo de planta en la que se encuentren. Además de agua, las vacuolas contienen típicamente sales y azúcares, y algunas proteínas en disolución. Debido al transporte activo y retención de ciertos iones por parte del tonoplasto, los iones se pueden acumular en el líquido vacuolar en concentraciones muy superiores a las del citoplasma exterior. A veces la concentración de un determinado material es suficientemente grande como para formar cristales, por ejemplo, de oxalato de calcio, que pueden adoptar distintas formas: drusa, con forma de estrellas, y rafidios, con forma de agujas. Algunas vacuolas son ácidas, como por ejemplo la de los cítricos. La vacuola, es a menudo un lugar de concentración de pigmentos. Los colores azul, violeta, púrpura, rojo de las células vegetales se deben, usualmente, a un grupo de pigmentos llamados antocianinas (responsables de las coloraciones de frutas y verduras).
- Funciones
Gracias al contenido vacuolar y al tamaño, la célula, el consumo de nitrógeno del citoplasma, consigue una gran superficie de contacto entre la fina capa del citoplasma y su entorno. El incremento del tamaño de la vacuola da como resultado también el incremento de la célula. Una consecuencia de esta estrategia es el desarrollo de una presión de turgencia, que permite mantener a la célula hidratada, y el mantenimiento de la rigidez del tejido, unas de las principales funciones de las vacuolas y cloroplasto. Otras de las funciones es la de la desintegración de macromoléculas y el reciclaje de sus componentes dentro de la célula. Todos los orgánulos celulares, ribosomas, mitocondrias y plastidios pueden ser depositados y degradados en las vacuolas. Debido a su gran actividad digestiva, son comparadas a los orgánulos de las células animales denominados lisosomas. También aíslan del resto del citoplasma productos secundarios tóxicos del metabolismo, como la nicotina (un alcaloide). Existen otras estructuras que se llaman también vacuolas pero cuya función es muy diferente:
- Vacuolas pulsátiles: éstas extraen el agua del citoplasma y la expulsan al exterior por transporte activo (osmosis) .
- Vacuolas digestivas: se produce la digestión de sustancias nutritivas, una vez digeridas pasan al interior de la célula y los productos de desecho son eliminados hacia el exterior de la célula.
- Vacuolas alimenticias: función nutritiva, forma a partir de la membrana celular y del retículo endoplasmático.
- Observación microscópica
En el microscopio fotónico se puede observar la célula vegetal, y en ella plastidios (cloroplastos, amiloplastos, etc.) y refiriéndose a la vacuola, no se puede divisar su membrana (tonoplasto), pero se deduce su ubicación porque se pueden ver las cristalizaciones (drusas y rafidios) de algunas sustancias que componen el jugo celular y también sirve como una fuerte sustancia que combate el virus del VIH.
- pared celular
La pared celular es una capa rígida que se localiza en el exterior de la membrana plasmática en las células de plantas, hongos, algas, bacterias y arqueas. La pared celular protege el contenido de la célula, y da rigidez a ésta, funciona como mediadora en todas las relaciones de la célula con el entorno y actúa como compartimiento celular. Además, en el caso de hongos y plantas, define la estructura y otorga soporte a los tejidos y muchas más partes de la célula. La pared celular se construye a partir de diversos materiales, dependiendo de la clase de organismo. En las plantas, la pared celular se compone, sobre todo, de un polímero de carbohidrato denominado celulosa, un polisacárido, y puede actuar también como almacén de carbohidratos para la célula. En las bacterias, la pared celular se compone de peptidoglicano. Entre las archaea se presentan paredes celulares con distintas composiciones químicas, incluyendo capas S de glicoproteínas, pseudopeptidoglicano o polisacáridos. Los hongos presentan paredes celulares de quitina, y las algas tienen típicamente paredes construidas a partir de glicoproteínas y polisacáridos. No obstante, algunas especies de algas pueden presentar una pared celular compuesta por dióxido de silicio. A menudo, se presentan otras moléculas accesorias integradas en la pared celular.
- Pared celular vegetal

- Estructura
La pared celular vegetal tiene tres partes fundamentales:
- Pared primaria. Está presente en todas las células vegetales, usualmente mide entre 100 y 200 nm de espesor y es producto de la acumulación de 3 o 4 capas sucesivas de microfibrillas de celulosa compuesta entre un 9 y un 25 % de celulosa. La pared primaria se crea en las células una vez que está terminando su división, generándose el fragmoplasto, una pared celular que dividirá a las dos células hijas. La pared primaria está adaptada al crecimiento celular. Las microfibrillas se deslizan entre ellas produciéndose una separación longitudinal, mientras el protoplasto hace presión sobre ellas.
- Pared secundaria. Cuando existe, es la capa adyacente a la membrana plasmática. Se forma en algunas células una vez que se ha detenido el crecimiento celular y se relaciona con la especialización de cada tipo celular. A diferencia de la pared primaria, contiene una alta proporción de celulosa, lignina o suberina.
- Laminilla media. Es el lugar que une las paredes primarias de dos células contiguas; es de naturaleza principalmente péptica, pero a menudo, en las células más viejas se lignifica.

- Composición
La composición de la pared celular vegetal varía en los diferentes tipos celulares y en los diferentes grupos taxonómicos. En términos generales la pared celular vegetal está compuesta por una red de carbohidratos, fosfolípidos y proteínas estructurales embebidos en una matriz gelatinosa compuesta por otros carbohidratos y proteínas.
- Carbohidratos
El principal componente de la pared celular vegetal es la celulosa. La celulosa es un polisacárido fibrilar que se organiza en microfibrillas y representa entre el 15 % y el 30 % del peso seco de las paredes celulares vegetales. Las microfibrillas de celulosa se encuentran atadas por carbohidratos no fibrilares a los que se denomina genéricamente hemicelulosa. Los componentes mayoritarios de la hemicelulosa son xiloglicanos (XiGs) glucuronarabinoxilanos (GAXs). La pectina es otro componente importante de las paredes celulares. Es un polisacárido no fibrilar, rico en ácido D-galacturónico, heterogéneamente ramificado y muy hidratado. Los componentes mayoritarios de la pectina son: los homogalacturonanos (HGA) y ramnogalacturonanos I (RG I). La matriz de pectina determina la porosidad de la pared y proporciona cargas que modulan el pH de la pared. Lignina y suberina son polímeros complejos compuestos por fenilpropanoides y alcoholes aromáticos. Se acumulan en algunas paredes secundarias y, en casos excepcionales, en paredes primarias. La lignina, la suberina y ceras como la cutina, le confieren impermeabilidad al agua a los tejidos en los que se depositan.
- Proteínas
La pared celular vegetal también está compuesta por proteínas estructurales. Estas proteínas son ricas en uno o dos aminoácidos, tienen dominios con secuencias repetidas y están glicosiladas en mayor o menor grado. Para la mayoría de las proteínas estructurales de la pared vegetal, se ha propuesto que tienen estructura fibrilar y que se inmovilizan mediante enlace entre ellas o con carbohidratos. Se sabe que estas proteínas se acumulan en la pared en diferentes etapas del desarrollo y en respuesta a diferentes condiciones de estrés. Se consideran proteínas estructurales de la pared celular vegetal: extensinas o proteínas ricas en hidroxiprolina (HRGPs), proteínas ricas en prolina (PRPs), proteínas ricas en glicina (GRPs) y arabinogalactanas (AGPs). Incluidas en la red de polisacáridos y proteínas, se encuentran diversas proteínas solubles, algunas de ellas son enzimas relacionadas con la producción de nutrientes como la glucosidasa, enzimas relacionadas con el metabolismo de la pared como las xiloglucano-transferasas, peroxidasas y lacasas, proteínas relacionadas con la defensa, proteínas de transporte.

- Biogénesis de la pared celular vegetal
La pared celular vegetal se constituye durante la división celular, a partir de vesículas que provienen del aparato de Golgi. Estas vesículas, llenas de los componentes de la pared celular, se localizan en el fragmoplasto, que es un arreglo del citoesqueleto propio de las células en división. En el fragmoplasto se fusionan las vesículas del aparato de Golgi y constituyen el plato celular el cual crece desde el interior de la célula en división, hasta ponerse en contacto con las paredes laterales. Una vez formada, la pared celular crece por deposición de capas sucesivas de celulosa. En cada capa, la orientación de las microfibrillas de celulosa está guiada por el citoesqueleto, más exactamente por los microtúbulos corticales, los cuales alinean al complejo responsable de la síntesis de celulosa, que es la celulosa sintasa. La elongación celular ocurre en el eje perpendicular al de las microfiblillas de la capa de pared que se está depositando, de ahí que la síntesis de la pared y la orientación de las microfiblillas de celulosa están en directa relación con el tamaño celular.

- Interacciones de la pared celular vegetal
La pared celular es el orgánulo más externo de la célula y de ella dependen las interacciones entre células y entre tejidos. Al igual que de la matriz extracelular de animales, de la pared celular de plantas depende la adhesión al substrato, la cual es determínate en el caso de algunas órganos vegetales que son móviles como el polen. De otro lado, la pared se mantiene en constante comunicación con el interior celular, esta interacción entre la pared y protoplasto es dinámica y transmite señales hacia el interior de la célula, que dan cuenta de las condiciones del ambiente extra-citoplasmático. En el otro sentido, de adentro hacia afuera, el protoplasto regula el estado de la pared en cada momento, dependiendo del desarrollo del tejido y las condiciones ambientales. Durante el fenómeno conocido como plasmólisis, que es la separación del protoplasto vivo de la pared celular por un efecto hiperosmótico, la interacción física entre la pared celular y el protoplasto se hace evidente; cuando esta interacción física se pierde la célula se vuelve incapaz de responder al ataque de patógenos y pierde su diferenciación celular.
- Pared celular de las algas
Al igual que las paredes celulares de las plantas superiores, la pared celular de las algas está compuesta por carbohidratos como la celulosa y glicoproteínas. La presencia de algunos polisacáridos en las paredes de las algas, es usada como carácter diagnóstico en la taxonomía de las algas.
- Polisacáridos sulfonados como la agarosa, se presentan en las paredes de algas rojas.
- Manosyl: Forma microfiblillas en la pared celular de algunas algas verdes de los géneros Codium, Dasycladus, Acetabularia, Porphyra y Bangia entre otros.
- Ácido alginílico: es un polisacárido común en la pared celular de las algas pardas.
El grupo de algas diatomeas sintetiza sus paredes celulares (también conocidas como frústulas o valvas) usando ácido silícico (específicamente ácido ortosilícico, H4SiO4). El ácido se polimeriza intracelularmente, y después la pared sale al exterior para proteger a la célula. Comparadas con las membranas celulares orgánicas producidas por otros grupos, requieren menos energía (aproximadamente el 8 %) para su síntesis, lo que constituye un ahorro para la célula, y posiblemente explique las tasas de crecimiento más altas en las diatomeas.
- Pared celular de los hongos
No todas las especies de hongos tienen paredes celulares, pero en el caso que las tengan, se componen de glucosamina y quitina, el mismo glúcido que da dureza a los exoesqueletos de los insectos. Tienen el mismo propósito que las paredes celulares de las plantas, dar rigidez a las células para mantener su forma y prevenir la lisis osmótica. También limita la entrada de moléculas que pueden ser tóxicas para hongo, tales como fungicidas sintéticos o producidos por plantas. La composición, las características y la forma de la pared celular de los hongos varían durante su ciclo vital y también depende de las condiciones de crecimiento. Oomycetes es un grupo de patógenos saprotróficos de las plantas que tienen paredes celulares anómalas de celulosa. Hasta hace poco tiempo se creía que eran hongos, pero datos estructurales y moleculares han llevado a su reclasificación como Heterokontophyta. Este es un grupo de protistas que incluye a autótrofos tales como algas pardas y diatomeas.
- Pared celular bacteriana

1-membrana citoplasmática
2-pared celular,
3-periplásmico. Abajo: Bacteria Gram negativa.
4-membrana citoplasmática,
5-pared celular,
6-membrana externa,
7-espacio periplásmico.

- En las bacterias Gram-positivas la pared celular contiene una capa gruesa de peptidoglicano además de ácidos teicoicos, que son polímeros de glicerol o ribitol fosfato. Los ácidos teicoicos se unen al peptidoglicano o a la membrana citoplasmática.
- En las bacterias Gram-negativas la capa de peptidoglicano es relativamente fina y se encuentra rodeada por a una segunda membrana lípida exterior que contiene lipopolisacáridos y lipoproteínas. La capa de peptidoglicano se une a la membrana externa por medio de lipoproteínas.
La mayoría de las bacterias tienen una pared celular Gram-negativa y solamente Firmicutes y Actinobacteria (conocidas previamente como bacterias Gram-positivas de contenido GC bajo y bacterias Gram-positivas de contenido GC alto, respectivamente) tienen paredes Gram-positivas. Estas diferencias en estructura pueden producir diferencias en la susceptibilidad antibiótica, por ejemplo, la vancomicina puede matar solamente a bacterias Gram-positivas y es ineficaz contra patógenos Gram-negativos, tales como Haemophilus influenzae o Pseudomonas aeruginosa.
- Pared celular de las arqueas
A diferencia de las bacterias, la pared celular arqueana no contiene peptidoglucano, en su lugar hay compuestos péptidos, glúcidos y/o glicoproteicos. Son de cuatro tipos:
- Pared de glicoproteínas, común en hipertermófilos, hiperhalófilos y metanógenos
- Pared de pseudopeptidoglicano, típica de las metanobacterias
- Capa S proteica, como en Methanomicrobia y Desulfurococcus
- Pared de polisacáridos, como en Methanosarcina y Halococcus
Algunas arqueas no poseen pared, como la clase Thermoplasmata.
- tráfico intracelular de membranas
Tráfico intracelular de membranas Los distintos compartimentos que componen las células eucariotas no son estructuras estáticas, sino todo lo contrario: son estructuras altamente organizadas y muy dinámicas. Así existe un flujo de compartimentos membranosos desde el interior de la célula hacia el exterior y viceversa. Los compartimentos celulares que intervienen en las rutas membranosas internas son:
- Retículo Endoplasmático (RE)
- VTC’s o ERGIC
- Aparato de Golgi o dictiosomas
- Endosomas:
- Endosoma temprano (EE, Early Endosome o Seaching Endosome)
- LE Late Endosome
- RE Recycling Endosome
- Lisosomas
- Caveosomas
- Notables biólogos celulares o citólogos
- Peter Agre
- Günter Blobel
- Christian de Duve
- Robert Hooke
- H. Robert Horvitz
- Anton van Leeuwenhoek
- Peter Dennis Mitchell
- Organización del cuerpo de la planta
El cuerpo de las plantas vasculares está marcadamente polarizado y formado por dos porciones básicas: un vástago orientado hacia la luz, que vive en ambiente aéreo, compuesto por tallo y hojas, y una raíz, órgano de fijación y absorción que vive en el suelo. Este tipo de cuerpo vegetativo se llama cormo y se presenta en las pteridofitas y en las espermatofitas, que por eso se llaman también cormófitos. Es difícil hacer una distinción entre tallo y hojas, ambos órganos tienen origen común en el meristemo apical caulinar, y están relacionados con estrecha dependencia a lo largo de todo su período de crecimiento. Por eso tallo y hojas se consideran como una unidad que constituye el vástago. En las espermatofitas la diferenciación entre raíz y vástago aparece ya en el embrión joven. Las partes del embrión son radícula, hipocótilo, cotiledones y plúmula. En algunos casos se distingue también el primer entrenudo, entre el nudo cotiledonar y la plúmula: el empicótelo. Durante la germinación el embrión crece, la radícula formará la raíz primaria y la plúmula formará el vástago.








